O tym jak biolog ewolucyjny doktor Bartosz Borczyk mówił o komputerowej „symulacji ewolucji oka” Nilssona i Pelgera, nie wiedząc o czym mówi!

Był czas, że wiedziałem o biologii mało. Był też taki że dość dużo, ale znacznie mniej niż obecnie. Teraz jest czas, że wiem mniej niż będe wiedział za tydzień, rok, czy pięć lat -o ile dożyję, bo w tym świecie więcej sposobów jest na umieranie niż na przeżywanie :) Człowiek uczy się całe życie! Jednak zawsze wychodziłem z założenia, że jak się chce krytykować jakąkolwiek koncepcję naukową, to najpierw należy się dokładnie zapoznać z przeciwną argumentacją. Niestety wielu biologów ewolucyjnych – za takich się przynajmniej podają – nie czuje potrzeby zapoznania się nie tylko z opiniami przeciwników, ale nawet z argumentacją czołowych biologow ewolucyjnych. Jednym z takich „fachowców” od ewolucji jest doktor Bartosz Borczyk:
Na powyższym filmie przeprowadza z nim wywiad niejaki pan Fjałkowski, który ma aspiracje zostania polskim Richardem Dawkinsem. Sam zaś doktor Bartosz Borczyk już nie jeden raz służył swoją (nie)wiedzą portalowi racjonalista.pl, gdzie w sposób jawnie niemerytoryczny usiłował podważać tezy Michaela Behe'ego - jego koncpecję nieredukowalnej złożoności. Podczas dyskusji panów Fjałkowskiego i Borczyka wyłonił się problem ewolucji złożonych organów. Doktor Borczyk od razu przywołał przykład oka, który z resztą mierzwił już samego Karola Darwina.
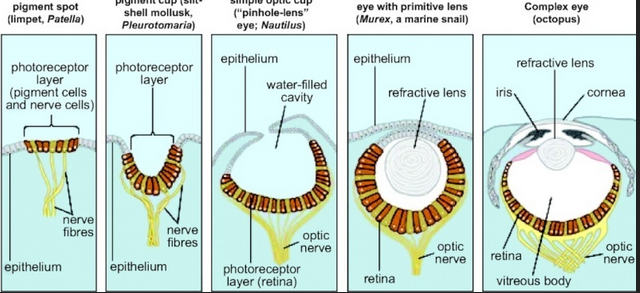
Następnie, żeby dowodzić możliwości ewolucji oka od komórek światłoczułych zaczynając, pan Borczyk przywołał pewien eksperyment prawie wcale nie wiedząc o czym mówi. Więc czego doktor Bartosz Borczyk nie potrafił nazwać po imieniu - dokładnie sprecyzować i o tym opowiedzieć, wiedząc że gdzieś tam gra, a nie wiedział dokładnie gdzie? Chodzi o niemal kultowy i znany każdemu porządnemu darwiniście eksperyment przeprowadzony przez Nilssona i Pelgera, który rzekomo symulował ewolucję oka na poziomie tkanek (fenotypu). Richard Dawkins dokładnie go opisał w swojej „Wspinaczce na szczyt nieprawdopodobieństwa” i zaprezentował w występach dla dzieci, które zostały zarejestrowane i zamieszczone na youtubie:
Rzekoma symulacja ewolucji oka była przeprowadzona na poziomie tkanek, a nie genów, czy procesów odpowiedzialnych za ich wyrażanie. Najpierw skoncentrujemy się na (nie)możliwości ewolucji komórek światłoczułych, bo to od nich miała się zacząć ewolucja oka, a następnie napiszę o reszcie. Fototransdukcja, to jeden z procesów, jakie opisal Michael Behe, uznając go za kompleks nieredukowalnie złożony:

Z punktu widzenia biochemii proces widzenia wygląda dość skomplikowanie:
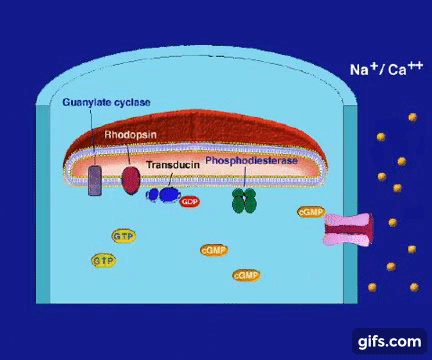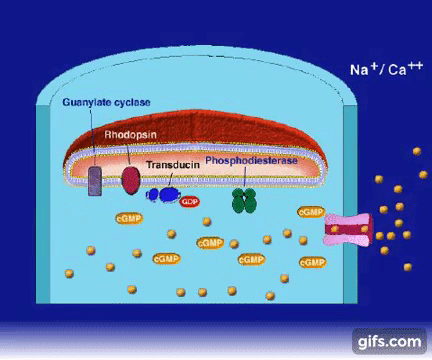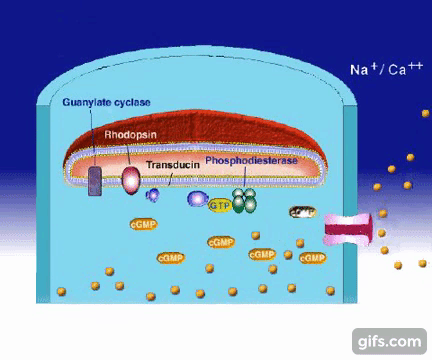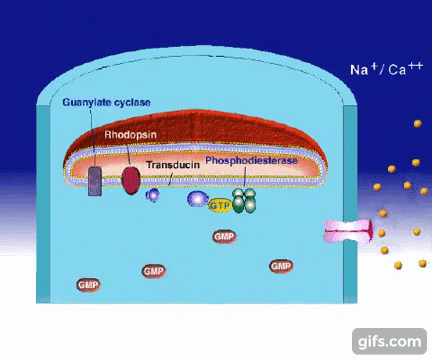
1.Gdy światło dociera do siatkówki, foton reaguje z cząsteczką (białkiem) zwaną 11-cis-retinalem, która w ciągu kilku pikosekund przemienia się w trans-retinal.
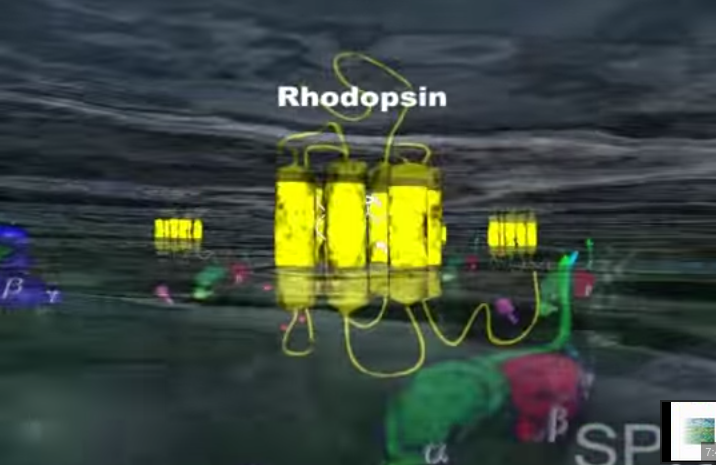

Zmiana kształtu (konformacji) cząsteczki retinalu wymusza zmianę kształtu białka- rodopsyny, z którym retinal jest ściśle związany. Z metamorfozą białka idzie w parze zmiana jego funkcji. Białko nazywane teraz metarodopsyną II przyczepia się do innego białka zwanego transducyną. Przed przylgnięciem do metarodopsyny II , transducyna mocno wiązała małą molekułę zwaną GDP. Lecz gdy transducyna reaguje z metarodopsyną II , GDP zostaje uwolniony, a z transducyną wiąże się molekuła GTP (GTP jest blisko spokrewnione z GDP, ale różnią się one istotnie).
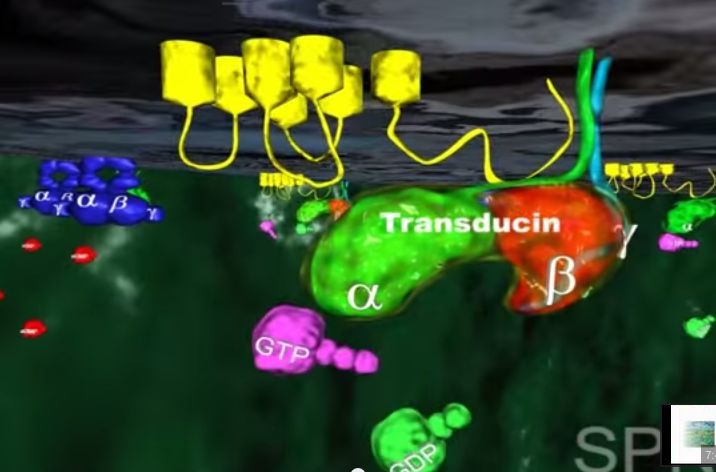
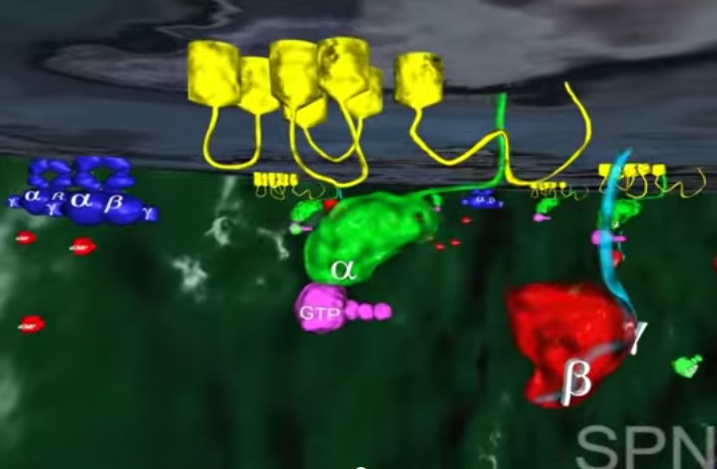
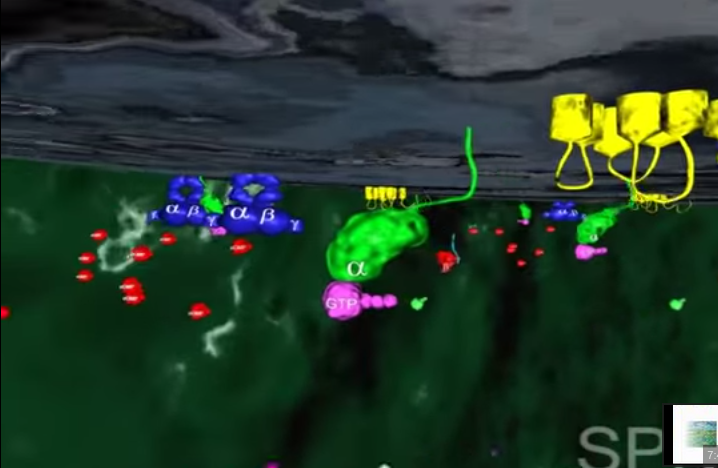
2.W tym momencie GTP-transducyno-metarodopsyna II wiąże się z białkiem zwanym fosfodiestrazą, które znajduje się w wewnętrznej błonie komórki. Po związaniu z metarodopsyną II i jej otoczeniem, fosfodiestraza nabywa zdolności „odcinania” cząsteczki zwanej cGMP. Początkowo w komórce znajduje się wiele cząsteczek cGMP, ale fosfodiestraza obniża ich koncentrację tak, jak wyjęcie korka powoduje obniżenie poziomu wody w wannie.
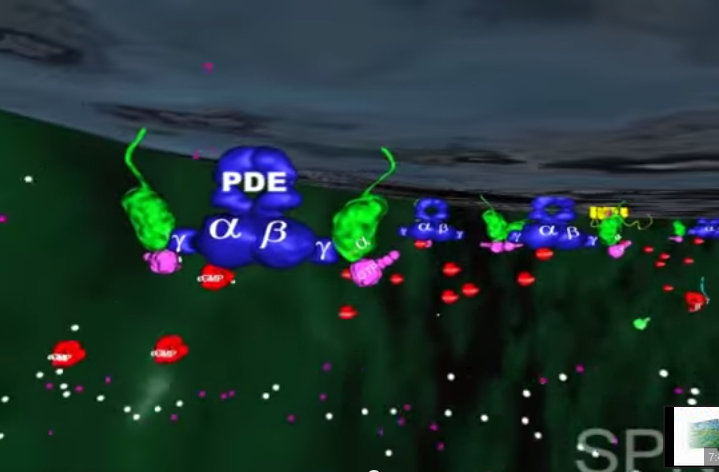
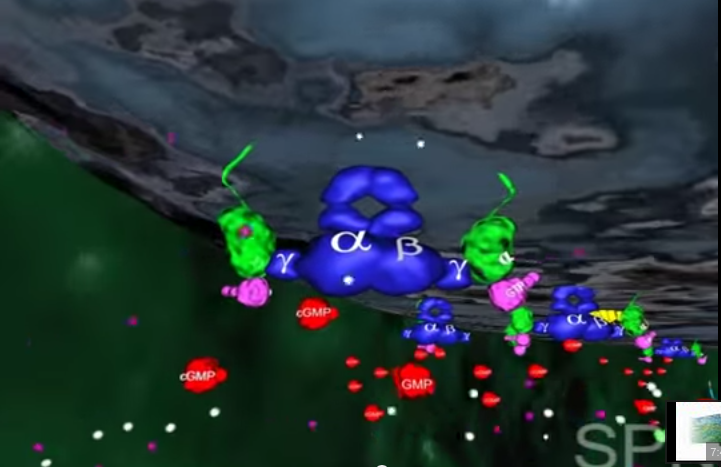
3.Inne białko błonowe, wiążące się z cGMP zwane jest kanałem jonowym. Działa on jak brama wjazdowa, która reaguje liczbę jonów sodu w komórce. Zwykle kanał jonowy pozwala na wpływanie jonów sodu do komórki, gdyż osobne białka aktywnie wypompowują je z powrotem. Wspólne działanie kanału jonowego i pompy utrzymuje w komórce w miarę stały poziom jonów sodu. Gdy ilość cGMP w komórce zostaje zredukowana z powodu rozczepienia przez fosfodiestrazę, kanał jonowy zamyka się powodując obniżenie dodatnio naładowanych jonów sodu w komórce.
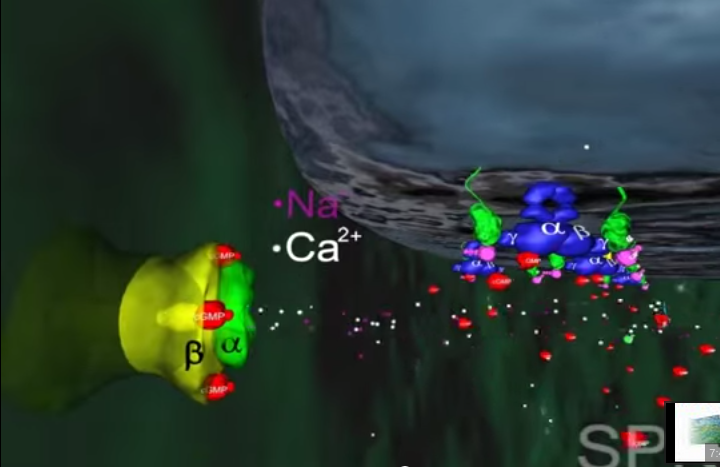
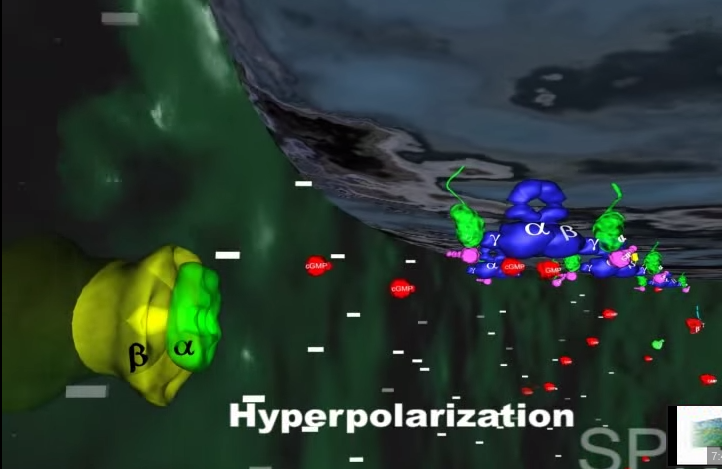
4.To z kolei powoduje zachwianie potencjału elektrycznego w komórce i wysyłanie impulsu do mózgu przez nerw wzrokowy. Efektem tego, po zinterpretowaniu przez mózg, jest widzenie.
Potencjał czynnościowy, czyli na czym polega impuls nerwowy
https://www.neuronaukowiec.com/potencjal-czynnosciowy/
https://pl.wikipedia.org/wiki/Potencja%C5%82_czynno%C5%9Bciowy
"Potencjał czynnościowy, potencjał iglicowy – przejściowa zmiana potencjału błonowego komórki, związana z przekazywaniem informacji. Bodźcem do powstania potencjału czynnościowego jest zmiana potencjału elektrycznego w środowisku zewnętrznym komórki. Wędrujący potencjał czynnościowy nazywany jest impulsem nerwowym."

5.Gdyby opisane wyżej reakcje były jedynymi zachodzącymi w komórce, to zasoby 11-cis-retinalu, cGMP i jonów sodu szybko by się wyczerpały. Coś (jakieś procesy) musi wyłączać białka, które były włączone i przywracać komórce jej stan pierwotny. Uczestniczy w tym kilka mechanizmów biochemicznych. Po pierwsze: w ciemnościach kanał jonowy oprócz jonów sodu wpuszcza jeszcze do komórki także jony wapnia. Są one wypompowywane z powrotem przez inne białko tak, żeby utrzymywała się stała koncentracja wapnia. Gdy opada poziom cGMP , za przyczyną zamknięcia kanału jonowego, obniża się również koncentracja jonów wapnia. Enzym fosfodiestraza, który niszczy GMP powodując spadek jego stężenia w komórce, spowalnia swoje działanie przy mniejszej koncentracji wapnia
6.Po drugie: gdy zaczyna się obniżać poziom wapnia, białko zwane cyklazą guanylową rozpoczyna ponowną syntezę cGMP.
7.Po trzecie: w trakcie tego procesu metarodopsyna II zostaje zmodyfikowana przez enzym zwany kinazą rodopsynową . Zmodyfikowana rodopsyna wiąże się wówczas z białkiem zwanym jako arestyna, które nie dopuszcza do aktywacji przez rodopsynę większej ilości transducyny
8.W komórce więc zachodzą reakcje zapobiegające wzmocnieniu sygnału wywołanego przez pojedynczy foton.
9.Trnas-retinal zostaje ostatecznie uwolniony z rodopsyny i musi ponownie być przemieniony w w 11-cis-retinal, następnie zostaje związany przez rodopsynę, wracając do punktu wyjściowego kolejnego cyklu procesu widzenia .
10.Aby to osiągnąć trans-retinal jest najpierw chemicznie przekształcany przez enzym w trans-retinol -formę posiadającą dwa dodatkowe atomy wodoru.
11.Drugi enzym następnie przekształca tą molekułę w 11-c-s-retinol.
12.W końcu trzeci enzym usuwa wcześniej dodane dwa atomy wodoru, tworząc 11-sic-retinal. Cykl się zamyka.
Rodopsyna jest białkiem fotoreceptorowym zlokalizowanym w komórkach pręcikowych siatkówki oka. Receptor ten bierze udział w kaskadach sygnalizacyjnych umożliwiających przekształcenie sygnału świetlnego w impuls nerwowy :
https://bioexploratorium.pl/wiki/Rodopsyna

Zawiłości rozwoju oka na poziomie genów. Czy geny odpowiedzialne za morfogenezę oka mogły wyewoluować stopniowo, po kolei za każdym razem je ulepszając jak w symulacji komputerowej Nilssona i Pelgera, a rzekome ewolucyjne prekursory oka zachowywać funkcję na każdym etapie ewolucji - dając przewagę selekcyjną?
Według scenariusza neodarwinowskiego ewolucji oka, komórki światłoczułe (u ssaków czopki i pręciki) wyewoluowały jako pierwsze. Jeżeli jednak założymy, że poszczególne etapy morfogenezy oka w jakiś sposób odzwierciedlają ewolucyjne powstawanie tego organu, to zauważymy, że mimo iż komórki światłoczułe rzeczywiście wykształcają się na początku rozwoju oka, to jednak niemal równocześnie z soczewką.
Upraszczanie do granic prostactwa!
Darwinowski scenariusz na podstawie symulacji ewolucji oka na poziomie tkanek przeprowadzonej przez Nilssona i Pelgera:

Poszczególne etapy rozwoju oka podczas morfogenezy tego organu:
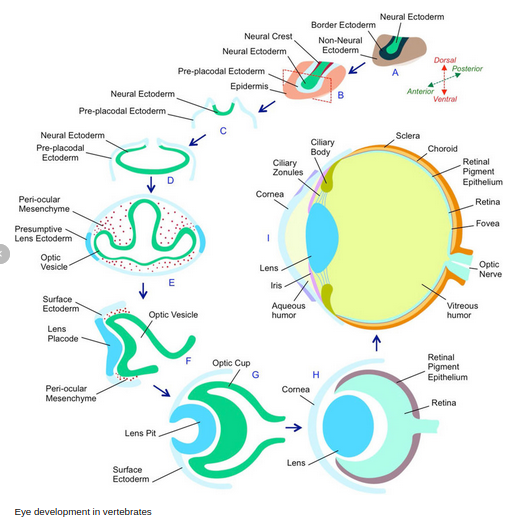
http://www.pum.edu.pl/__data/assets/file/0008/57995/WLA-I-ROK-Zmysly-2013.pdf (strony 26, 27 i 28).
1) Zagłębienia w cewie nerwowej (przyszłe przodomózgowie) – rowki wzrokowe
2) Pęcherzyki wzrokowe (po zamknięciu cewy nerwowej) na zewnętrznej powierzchni
zawiązka CUN
3) Koniec 4 tygodnia – światło pęcherzyka szerokie i łączy się ze światłem przedmózgowia
4) Początek 5 tygodnia – odcinek dystalny pęcherzyka wpukla się – kubek wzrokowy o podwójnej ścianie
5) Intensywne różnicowanie obu ścian. W-wa zewnętrzna cieńsza , w 6 tygodniu – melanina (warstwa barwnikowa siatkówki). W-wa wewnętrzna grubieje – wzrokowa część siatkówki (blaszka nerwowa).
6) Różnicowanie w polu środkowym ku obwodowi.
– Spongioblasy-komórki tkanki glejowej
– Neuroblasty – komórki nerwowe siatkówki
Najpóźniej ok. 7 m-ca życia ostatecznemu różnicowaniu ulegają komórki pręcikonośne
i czopkonośne.
Soczewka
1) Wraz z kubkiem wzrokowym – zawiązek soczewki.
2) 4/5 tydzień rozwoju – ektoderma pokrywowa – płytka soczewki, zagłębiając się do
światła kubka – pęcherzyk soczewki.
3) 5 tydzień – pęcherzyk soczewki zamyka się i oddziela się od ektodermy.
4) Koniec 6 tygodnia – komórki tylnego bieguna – włókna soczewki, wypełniają wnętrze
pęcherzyka soczewki.
5) Komórki bieguna przedniego – w-wa komórek sześciennych.
6) Komórki okolicy równikowej – zachowana zdolność do podziału (warstwa rozrodcza) – wzrost przez apozycję.
7) Brak własnych naczyń, odżywiana przez naczynia otaczające.
Ciało szkliste
8)Komórki zrębu siatkówki wysyłają cienkie wypustki.
9) Włókienka + galaretowata wydzielina komórek siatkówki – ciałko szkliste
10) Tętnica centralna siatkówki – tętnice ciała szklistego.
Jak więc widzimy prekursorowa soczewka, podczas morfogenezy oka, zaczyna się formować niemal równocześnie z siatkówką, to znaczy z receptorami światłoczułymi. Następnie siatkówka, a za nią soczewka, wpukla się do zagłębienia (oczodołu) w ciele. Soczewka jest wówczas niefunkcjonalna, występuje w postaci pęcherzyka i zaczyna się dopiero rozwijać do formy funkcjonalnej, kiedy siatkówka ‚zagnieździ’ się już na dnie oka.
Zalążek soczewki, pęcherzyk soczewkowy, rozwija się niemal równocześnie z prekursorem siatkówki i na tym etapie, i jeszcze przez jakiś czas rozwoju oka nie nadaje się do tego, aby brać udział w procesie widzenia i żadne uciskanie jej, jak torebki z wodą, której użył Richard Dawkins nic tutaj nie pomoże. Soczewka nadaje się do widzenia dopiero wówczas, kiedy oko się niemal całkowicie rozwinie. Podczas zakładanej ewolucji oka taka nieprzezroczysta soczewka zasłoniłaby komórki światłoczułe i już na tym etapie ewolucja oka utknęłaby w martwym punkcie. Hipotetycznie ewolucja biologiczna opiera się na procesie selekcyjnego wymiatania przy udziale doboru kumulatywnego.
W tym przypadku prekursory oka musiały by dawać przewagę selekcyjną na każdym etapie wiodącym do budowy organu, jaki istnieje obecnie. W komputerowej symulacji Nilssona i Pelgera badacze założyli, że przewagę daje coraz lepsze, ostrzejsze widzenie, co pozwalało na sprawniejsze zdobywanie pożywienia i unikanie drapieżników i innych niebezpieczeństw.W kolejnych pokoleniach coraz lepiej widzące organizmy okazywały się sprawniejsze od tych, które nie nabyły odpowiednich modyfikacji ewolucyjnych w postaci mutacji, które by powstały w gametach [komórkach rozrodczych] ich rodziców, lub które by odziedziczyli po nich.
Szybciej niż oni zdobywali pożywienie, sprawniej unikali drapieżników i w efekcie organizmy te przeżywały wydając większą liczbę potomstwa. Z czasem organizmy te oraz ich potomstwo, które odziedziczyło cechy gwarantujące przewagę nad tymi, którzy nie mieli szczęścia zdobyć, lub odziedziczyć potrzebnych mutacji eliminowało [wymiatało] konkurencję. Soczewka, która działałaby jak zasłona nie pomogłaby w walce o przetrwanie. Załóżmy, że ludzkość zaczęła między sobą bezwzględną wojnę na śmierć i życie o kurczące się zasoby. Nagle w wyniku promieniowania kosmicznego 99% ludzkiej populacji traci wzrok. Kto by miał największe szanse na wygranie tej wojny, zdobycie pożywienia i pozostawienie potomstwa?
Geny biorące udział w rozwoju oka działają na kanwie ścisłej współpracy i na zasadzie plejotropii;
http://pl.wikipedia.org/wiki/Plejotropia .
Ich współpraca odbywa się na tej zasadzie, że te same geny regulują różne procesy, na różnych etapach w rozwoju oka biorąc udział w różnych wzorach ekspresji genów. Oto jak PZ Myers, profesor biologii i ewolucjonista, tłumaczy tą genetyczną aktywność w rozwoju oka:
file:///tmp/mozilla_slawek0/racjonalista_7994.pdf
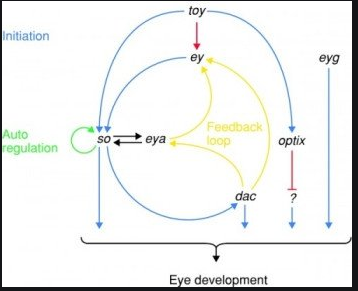
„ Schemat regulatorowy na szczycie szlaku rozwojowego oka. Twin of eyeless (toy),eyeless (ey), i być może eyegone (eyg), trzy geny Pax, są głównymi genami kontrolującymi na szczycie hierarchii. Sine oculis (so), eyes absent (eya), dachshund (dac), i optix są drugorzędowymi czynnikami transkrypcyjnymi, regulowanymi przez główne geny kontrolne. Proszę zauważyć, że szlak nie jest liniowy, ale raczej jest siecią powiązaną przez pętle sprzężenia zwrotnego.
Na szczycie hierarchii są u Drosophila dwa geny: eyeless (ey) i twin of eyeless (toy). Pamiętajmy, że geny są nazwane od efektów mutantów, a więc normalną funkcją eyeless jest zainicjowanie rozwoju oka. Te geny włączają sine oculis i eyes absent (proszę zauważyć wysiłek znalezienia synonimów, żeby opisać geny, których brak lub złe działanie powoduje brak oczu), które aktywują się wzajemnie i zwracają sygnał do eyeless, żeby stworzyć silną reakcję. Inny gen, dachshund (ten gen jest nazwany od innej części jego fenotypu: tworzy muchy z bardzo krótkimi nogami) także zwraca sygnał do eyeless.
Ten obwód ma wiele wyjść: so, dac, optix and eyg. Wszystkie one wywierają efekty w procesie określanym szeroko jako „rozwój oka”. W tej szerokiej kategorii leży wiele procesów: wyżej wspomniany szlak pigmentacji, ale także wszelkiego rodzaju wyrafinowane interakcje, które rekrutują komórki do specjalnych funkcji fotoreceptorów, które organizują komórki pomocnicze, jak komórki włoskowe i soczewki, i które indukują tkankę nerwową siatkówki i głębszych części układu nerwowego. Geny ey i toy inicjują całą głęboką, rozgałęziającą się sieć genów, które razem, w kaskadzie, budują wiele części składowych oka.
Te dwa główne geny kontrolne, eyeless i twin of eyeless, mają także synonim. Ku zaskoczeniu wszystkich wersja tego obwodu znajduje się u wszystkich zwierząt mających oczy, a powszechną nazwą tego uniwersalnego regulatora tworzenia oka jest Pax6. [….]„
W świetle tych faktów zadziwia, że darwiniści nie posiadając modelu teoretycznego, który chociaż pobieżnie opisywałby ewolucję najprostszego oka twierdzą, że oko wyewoluowało 40 do 60 razy niezależnie w historii ziemi! :)
http://eyedesignbook.com/
Homologiczne geny kodują niehomologiczne struktury?
Oko ośmiornicy jest bardzo podobne morfologicznie do oka ssaków (człowieka). 729 wspólnych sekwencji u ośmiornicy i człowieka z 1052 przebadanych genów, to 70% wspólnych genów z człowiekiem. Aby prześledzić możliwość zmian ewolucyjnych, które były potencjalnie odpowiedzialne za ewolucję oczu u ośmiornic i innych organizmów, przeprowadzono analizę porównawczą. Okazało się, że 1019 spośród 1052 genów biorących udział w rozwoju oczu jest wspólna dla różnych organizmów, u których rzekomo niezależnie wyewoluowały oczy. Jedne z tych organizmów mają mniej podobnych genów do innych gatunków z oczami, drugie mają ich więcej. Patrząc ewolucyjnie te geny musiałyby istnieć już u wspólnego przodka różnych gatunków z oczami, ale tak być nie mogło, ponieważ inne geny nie są homologiczne. Zatem skąd one się tam wzięły? Jak to się stało że homologiczne z punktu widzenia darwinistów geny kodują niehomologiczne oczy, te które miały powstać niezależnie kilkadziesiąt razy w historii ziemi? Oto wielka tajemnica wiary neodarwinizmu!!
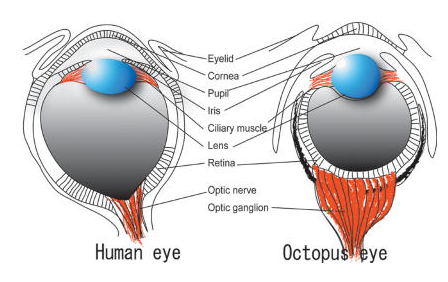
Problem istnienia tzw. głębokiej homologii, bo tak darwiniści nazwali zjawisko kodowania niehomologicznych organów (które nie mogły powstać od wspólnego przodka) przez homologiczne geny, które z ich punktu widzenia muszą pochodzić od wspólnego przodka jest w biologii powszechny!
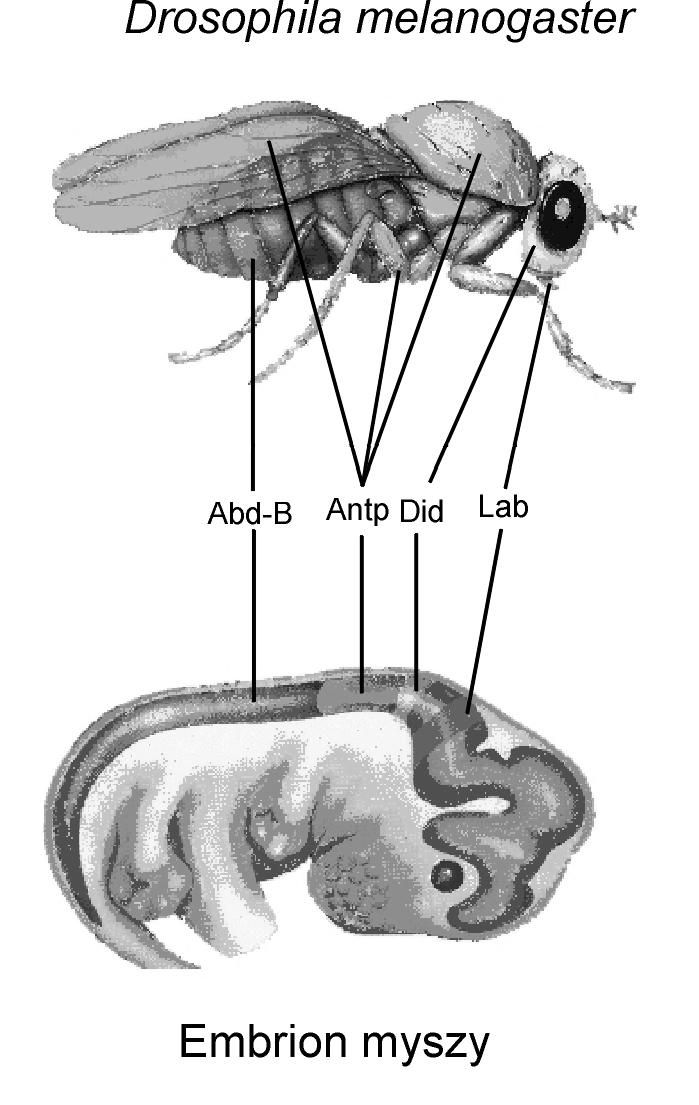
"Homologiczne" geny homeotyczne (Hom i Hox) występują u muszek owocowych i u myszy. Geny te — dziedziczone jakoby od wspólnego przodka, czyli homologiczne — sterują rozwojem niehomologicznych struktur.
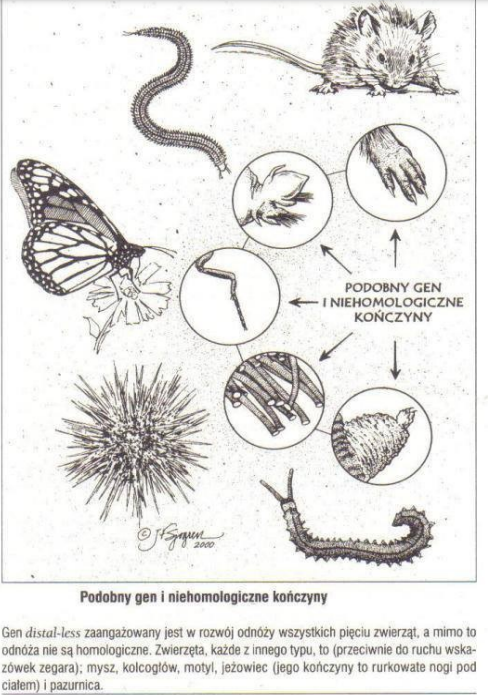
Brak związków pomiędzy genami i strukturami dotyczy nie tylko całych struktur, ale i poszczególnych organów. Np. oczu, czy kończyn. Gen distal – less, to gen sterujący rozwojem kończyn u wielu różnych typów zwierząt. I mimo że te kończyny są z punktu widzenia teorii ewolucji niehomologiczne, to jednak z tego samego punktu widzenia gen distal – less u tych wszystkich typów jest genem homologicznym! Gen o prawie identycznej budowie, co gen distal – less został znaleziony u jeżowców, kolcogłowów (należących do typu dżdżownic) oraz należących do zupełnie innego typu pazurnic. Biolodzy, którzy w latach 90 dokonali tego odkrycia, jak Gregory Waj, stwierdzili, że jest to zjawisko trudne do wytłumaczenia [na kanwie teorii ewolucji]. Poza tym odkryto, że nie tylko gen distal – less steruje rozwojem tych niehomologicznych kończyn, ale całe sieci zaangażowane w rozwój tych niehomologicznych kończyn są podobne, a nawet identyczne. Mamy w tym przypadku do czynienia z paradoksem. Anatomia tych kończyn przemawia za ich niehomologicznością, czyli za tym, że nie było między nimi żadnej ciągłości ewolucyjnej. Natomiast ogromne podobieństwo genów biorących udział w morfogenezie tych struktur, patrząc ewolucyjnie, prowadzi do wniosku o takiej ciągłości.
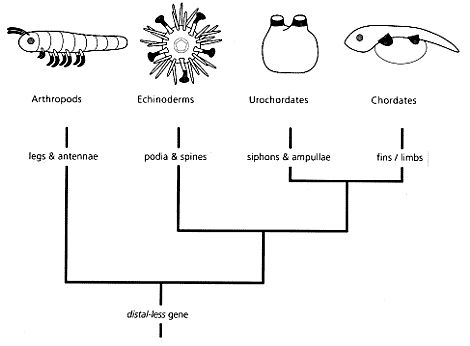
Gen 'distal-less' jest podobnie wyrażany w strukturach anatomicznych zaczernionych na diagramach (Wray in Briggs & Crowther 2001). Dzięki wiedzy o współczesnych i kopalnych taksonach wiadomo, że części te nie są homologiczne. Działanie genu distal-less nie zostało jeszcze poznane. Dane paleontologiczne dowodzą, że kończyny Tetrapoda są homologiczne płetwom ryb, a nie utworowi gębowemu osłonic Urochordata.
„(....)Oczy istnieją w różnych kształtach, rozmiarach, optycznych planach, różnią się położeniem w ciele, ale wszystkie ich rodzaje dostarczają ich posiadaczom podobnych informacji o długości fali i intensywności światła. Wśród różnych typów zwierząt do budowy soczewki i siatkówki wymagane są różne tkanki. Dla kontrastu, wszystkie rodzaje oczu posiadają ten sam mechanizm absorbowania fotonów, tj. kombinację opsynowo-chromoforową. Wspólny projekt oczu przejawia się więc w zastosowaniu podobnych „materiałów”, do budowy całej gamy różnego typu oczu. Co więcej, ostatnie badania donoszą, że bardzo zróżnicowane typy oczu formują się w bardzo podobnych rozwojowych procesach.
Rozwojem bardzo różnych typów oczu steruje u większości organizmów ten sam gen – Pax-6. Bardzo podobne są również kaskady genetycznych procesów, które jednak wiodą do zupełnie odmiennych końcowych rezultatów. Niektórzy ewolucjoniści argumentują w związku z tym, że oko ma genezę monofiletyczną. Większość biologów odrzuca jednak tę sugestię. Twierdzą oni, że po pierwsze są świadectwa, że ekspresja homologicznych genów Pax-6 zachodzi również w jeżowcach (Echinoidea) i nicieniach (Nematoda), które w ogóle nie posiadają oczu. Inne dowody wskazują, że gen ten uczestniczy w powstawaniu różnych tkanek mózgowych i jego aktywność nie jest ograniczona wyłącznie do rozwoju oka.
Co sprawia, że te same geny, te same rozwojowe procesy prowadzą do wykształcania się odmiennych funkcjonalnych rozwiązań? Homologiczne geny i mechanizmy rozwojowe prowadzą np. do budowy niehomologicznych oczu u zwierząt klasyfikowanych od siebie bardzo odlegle: kręgowców i stawonogów. Oczy kręgowców, funkcjonujące na zasadzie ciemni optycznej, zbudowane są na zupełnie odmiennej zasadzie niż oczy stawonogów (Arthopoda), które składają się z licznych (od kilkunastu do ok. 30 tys.) oczek prostych (ommatidiów) stanowiących pojedyncze układy optyczne; w oczach tego typu powstaje obraz mozaikowy (punktowy). Jednak, gdy gen Pax-6 myszy wszczepiony zostaje muszce owocowej (Drosophila) w trakcie jej embrionalnego rozwoju, jest on „interpretowany” właściwie – rozwija u niej normalne wielofasetkowe oko. Tak więc ten sam gen może „znaczyć” dwie różne rzeczy. U muszki gen ten uczestniczy w rozwoju oka fasetkowego, u myszy ten sam gen uczestniczy w rozwoju oka soczewkowego. Co więcej, ten sam gen zaangażowany u kręgowców w rozwój oka może również wywoływać rozwój... wizerunku oka na skrzydłach niektórych gatunków motyli. 28 Russell Fernald ze Stanford University podsumowuje:
Pomimo nowych danych dostarczanych przez potężne techniki molekularne, dostępne świadectwo wciąż sugeruje, że oczy mają polifiletyczne pochodzenie z zastrzeżeniem, że zawierają one homologiczne molekuły odpowiedzialne za wiele strukturalnych, funkcjonalnych i innych cech rozwojowych. Poznając wzrastającą liczbę homologicznych sekwencji genów pośród molekuł oka, wśród organizmów filogenetycznie bardzo odległych, aktualnym wyzwaniem jest odkrycie, co sprawia, że oczy Drosophila, kałamarnic i myszy są tak różne. Ponieważ ściśle homologiczne rozwojowe procesy muszą produkować homologiczne struktury, kluczowy element odpowiedzialny za rozwój niehomologicznych oczu wciąż
jest nieznany. Zrozumienie tego, co czyni oczy różnymi może być większym wyzwaniem, niż znajdowanie tego, co mają wspólnego.
Geny i procesy rozwojowe sterujące powstawaniem oczu zwierząt z fundamentalnie różnych grup: kręgowców, głowonogów i stawonogów, są w 94 procentach te same(....)”
Jednak darwiniści nie widzą problemu. Im więcej faktów przeczących filogenezie, tym gorzej dla faktów. "Głęboka homologia" musiała jakoś wyewoluować!! 'Wspinaczka na szczyt nieprawdopodobieństwa', Richard Dawkins napisał:

„(....) Mogłoby się wydawać, że twierdzeniu temu przeczą intrygujące wyniki eksperymentów, jakie przeprowadzili ostatnio w Szwajcarii naukowcy pracujący pod kierownictwem Waltera Gehringa. Przedstawię je pokrótce i udowodnię, że w istocie wcale nie są sprzeczne z naszą konkluzją. Zanim rozpocznę, muszę przeprosić czytelników za zwariowaną konwencję nazywania genów przez genetyków. Gen nazwany eyeless (po polsku - bezoki) u muszki owocowej Drosophila w istocie odpowiedzialny jest za powstanie oczu (prześliczne, prawda?!)! Powód tego figlarnie mylącego przykładu terminologicznej sprzeczności jest w rzeczywistości Richard Dawkinsdość prosty, a nawet interesujący. Otóż funkcję, jaką pełni określony gen, można rozpoznać wówczas, gdy dzieje się z nim coś złego. Kiedy gen, o którym mówimy, psuje się (czyli mutuje), u muchy nie rozwijają się oczy. Miejsce na chromosomie, w którym znajduje się ten gen, nazywane jest więc eyeless locus (locus to po łacinie miejsce, genetycy określają w ten sposób miejsce na chromosomie, w którym zlokalizowane są różne formy jednego genu). Mówiąc jednak o genie eyeless, mamy zwykle na myśli jego normalną, nieuszkodzoną wersję znajdującą się w tym locus. Stąd paradoks, że gen „bezoki” decyduje o powstawaniu oczu. To tak, jakby głośnik nazwać „wyciszaczem”, kiedy odkryje się, że po jego odłączeniu radio cichnie. Już przestaję. Mam tylko ochotę nazwać ten gen „eyemaker (po polsku - „okorób”), ale to chyba też specjalnie nie ułatwiłoby sprawy. Nie mogę jednak nazywać go eyeless, posłużę się więc skrótem ey.
Jak powszechnie wiadomo, choć wszystkie komórki każdego organizmu zawierają wszystkie geny, tylko niewielka ich część jest wykorzystywana (ulega ekspresji) w poszczególnych częściach ciała. Dlatego właśnie wątroba różni się od nerek, chociaż ich komórki zawierają dokładnie takie same geny. U dorosłej muszki owocowej gen ey zwykle ulega ekspresji tylko w komórkach głowy - dlatego oczy rozwijają się właśnie tam. George Haider, Patrick Callaerts i Walter Gehring opracowali procedurę powodującą powstawanie oczu także w innych rejonach ciała tego owada. Hodując larwy muszek w specjalny sposób, doprowadzili do tego, że gen ey uległ ekspresji w miejscach, w których normalnie powstają czułki, skrzydła i odnóża. Co jeszcze zabawniejsze, uzyskali dorosłe muchy z całkowicie rozwiniętymi oczami złożonymi na skrzydłach, odnóżach, czułkach i w innych miejscach (ryc. 5.29). Choć nieco mniejsze, oczy te mają całkiem normalną budowę i składają się z wielu właściwie ukształtowanych ommatidiów. A nawet działają. Nie wiemy, co prawda, czy muchy rzeczywiście coś przez nie widzą, ale rejestracja aktywności elektrycznej nerwów leżących u podstawy ommatidiów dowodzi, że są one wrażliwe na światło.
To jest godny uwagi fakt numer jeden. Fakt numer dwa jest jeszcze bardziej interesujący. Znany jest gen mysi, zwany small eye (po polsku - drobnooki) i gen ludzki, zwany aniridia (po polsku - bez tęczówki). Ich nazwy są także odzwierciedleniem negatywnej konwencji przyjętej przez genetyków: mutacyjne uszkodzenie tych genów powoduje zmniejszenie bądź brak oczu lub ich części. Rebecca Quiring i Uwe Waldorf z tego samego szwajcarskiego laboratorium stwierdzili, że oba te geny ssaków są niemal identyczne pod względem sekwencji DNA z genem ey muszki owocowej. Oznacza to, że ten sam gen przeszedł cały łańcuch przodków aż do obecnie żyjących zwierząt tak daleko ze sobą spokrewnionych jak owady i ssaki. Co więcej, u obu tych bardzo odległych grup zwierząt gen ten ma decydujące znaczenie w rozwoju oczu. Fakt numer trzy jest wręcz zdumiewający. Haiderowi, Callaertsowi i Gehringowi udało się wprowadzić gen mysi do zarodków muszek owocowych. Ku wielkiemu zdumieniu okazało się, że spowodował powstanie ektopowych oczu u Drosophila. Rycina 5.29 przedstawia niewielkie oczy złożone, jakie rozwinęły się na odnóżu muszki w wyniku indukcji mysim odpowiednikiem genu ey. Warto przy tym zwrócić uwagę, że powstało wówczas owadzie oko złożone, a nie oko charakterystyczne dla myszy. Mysi gen po prostu uruchomił mechanizmy embriogenetyczne odpowiedzialne za powstawanie oczu u muszki owocowej. Geny o niemal takiej samej sekwencji DNA jak gen ey znaleziono także u mięczaków, morskich robaków zwanych wstężnicami i u strzykw. Być może okaże się, że ey występuje u wszystkich zwierząt i podanie pewnej jego wersji pochodzącej z jednej grupy zwierząt przedstawicielowi jakiejś innej, bardzo odległej grupy spowoduje u niego rozwój oczu. Richard Dawkinsinduktorem oczu widocznych na najniższym zdjęciu był gen mysi.
Co wyniki tych spektakularnych doświadczeń znaczą dla konkluzji tego rozdziału? Czy myliliśmy się mówiąc, że oczy wykształciły się czterdzieści razy niezależnie od siebie? Nie sądzę. W mocy pozostaje przynajmniej duch twierdzenia, że oczy ewoluują łatwo i szybko. Wyniki te świadczą natomiast prawdopodobnie o tym, że wspólny przodek muszek owocowych, myszy, ludzi, strzykw i innych stworzeń miał oczy. Ten odległy przodek dysponował zapewne jakimś rodzajem wzroku, a o powstaniu jego oczu, niezależnie od ich postaci, prawdopodobnie zadecydowała sekwencja DNA podobna do tej, jaką ma dzisiejszy gen ey. Współczesne postaci różnego rodzaju oczu, szczegóły budowy ich siatkówek, soczewek czy zwierciadeł, wybór między okiem prostym a złożonym oraz między okiem apozycyjnym a różnymi odmianami oka superpozycyjnego są jednak rezultatem niezależnej i szybkiej ewolucji. Możemy się o tym przekonać, obserwując przypadkowe - można wręcz powiedzieć: kapryśne - rozrzucenie najróżniejszych rozwiązań, pomysłów i systemów w całym królestwie zwierząt. Mówiąc krótko: często badane zwierzę ma oczy przypominające dużo bardziej oczy swego bardzo odległego krewnego niż bliskiego kuzyna. Konkluzji tej w niczym nie narusza konstatacja, że wspólny przodek wszystkich zwierząt najprawdopodobniej miał jakieś oczy, i że rozwój zarodkowy oczu wszystkich zwierząt ma ze sobą wystarczająco dużo wspólnego, by można go było zaindukować tą samą sekwencją DNA. (….)”
Richard Dawkins o eksperymencie symulacji ewolucji oka Nilssona i Pelgera - polskie napisy:
Powszechne zwiedzenie!
Wyniki komputerowej symulacji ewolucji oka Nilssona i Pelgera, które z coraz większym rozmachem w kolejnych książkach opisał i odpowiednio przystroił w rzekomo naukowe piórka Richard Dawkins stanowią standardowy argument zwolenników neodarwinizmu. Na bezrybiu i rak ryba! :)
http://www.biologianaukaozyciu.pl/2017/07/ewolucja-oka.html

Poniższy fragment dłuższego wywodu nie stanowi rzetelnego przekazu opartego na wiedzy naukowej, tylko jest kolejną wersją pozornych mądrości Richarda Dawkinsa zaczerpniętą z książki popularnonaukowej o znamionach fantasy. Do udzielania korepetycji się nie nadaje, tylko do robienia wody z mózgów! :)

Napisałem, że książki Richarda Dawkinsa mają znamiona fantasy, ale on sobie lubi pochlebiać przekonując, że posiadają wyższy status!
https://doci.pl/Beatrycze99/dawkins-richard-samolubny-gen+fcvc51



O tym ile naukowości jest w książce Samolubny gen, której treść owładnęła umysły biologów niemal z tą samą mocą, co książka Karola Darwina O pochodzeniu gatunków na drodze doboru naturalnego - zanim obie koncepcje zostały zweryfikowane - opowiedział profesor Paweł Golik. Jeżeli ktoś potrafi odsiać pszenicę od plew - prawdę naukową od tego, co panu Golikowi się wydaje, albo od tego co chciałby uznawać za prawdę, to tylko zyska słuchając jego wywodu:
https://www.salon24.pl/u/adaptacjeslawekp/1117012,samolubny-gen-prf-richarda-dawkinsa-czy-samolubne-geny-prf-pawla-golika


Paweł Poręba, to najbardziej pyskaty uczestnik naszych dawnych dyskusji - biolog, zwolennik darwinizmu. Z 25 lat temu był postrachem kreacjonistów. Dość duża wiedza, pasja, pomysłowość oraz bezlitosny i twardy przekaz. Miałem okazję porozmawiać z nim drogą e-mailową tuż przed definitywnym wycofaniem się Pawła z internetowych bojów między darwinistami i kreacjonistami. Dyskutowaliśmy o (nie)możliwości stopniowego wyewoluowania przeobrażenia zupełnego (holometabolii) u owadów: https://www.salon24.pl/u/adaptacjeslawekp/1116580,od-gasienicy-do-motyla-cudowny-proces-holometabolii
Pamiętam że zadawałem mu różne trudne pytania, a on nie potrafił znaleźć wyczerpujących wyjaśnień. W końcu temat się wyczerpał, Paweł opowiedział trochę o nurkowaniu i temat się się pożegnaliśmy.
Wybrał karierę płetwonurka, napisał mi że to jego największa pasja obok biologii. Z czasem całkowicie się jej oddał. Paweł to gość z charakterem, co udowodnił podczas akcji ratowniczej swoich kolegów, którym śmiało poszedł na ratunek ryzykując życiem podczas awarii w konińskiej elektrowni. Tutaj jest jego sprawozdanie z tej akcji: https://www.nurkomania.pl/a0240.htm
Wykład Pawła Poręby na temat nurkowania technicznego w ramach Czwartkowych Spotkań w Divemanii
https://vimeo.com/20369307

https://ebookpoint.pl/ksiazki/czarna-skrzynka-darwina-biochemiczne-wyzwanie-dla-ewolucjonizmu-michael-j-behe,e_1um0.htm

Paweł Poręba, to najbardziej pyskaty uczestnik naszych dawnych dyskusji – biolog, zwolennik darwinizmu. Z 25 lat temu był postrachem kreacjonistów. Dość duża wiedza, pasja, pomysłowość oraz bezlitosny i twardy przekaz. Miałem okazję porozmawiać z nim drogą e-mailową tuż przed definitywnym wycofaniem się Pawła z internetowych bojów między darwinistami i kreacjonistami. Dyskutowaliśmy o (nie)możliwości stopniowego wyewoluowania przeobrażenia zupełnego (holometabolii) u owadów: https://www.salon24.pl/u/adaptacjeslawekp/1116580,od-gasienicy-do-motyla-cudowny-proces-holometabolii
Pamiętam że zadawałem mu różne trudne pytania, a on nie potrafił znaleźć wyczerpujących wyjaśnień – w końcu nie jest entomologiem i słusznie zauważył, że biolog nie musi się znać na każdej dziedzinie w obrębie nauk przyrodniczych. W końcu temat się wyczerpał. Paweł opowiedział trochę o nurkowaniu i się się pożegnaliśmy. Brakuje takiego dyskutanta we współczesnej sieci- istna maszyna polemiczna. Czasy świetności USENETU przeminęły ????
Paweł wybrał karierę płetwonurka, napisał mi że to jego największa pasja obok biologii. Z czasem całkowicie się jej oddał. Paweł to gość z charakterem, co udowodnił podczas akcji ratowniczej swoich kolegów, którym śmiało poszedł na ratunek ryzykując życiem podczas awarii w konińskiej elektrowni. Tutaj jest jego sprawozdanie z tej akcji: https://www.nurkomania.pl/a0240.htm
Wykład Pawła Poręby na temat nurkowania technicznego w ramach Czwartkowych Spotkań w Divemanii
http://www.deltami.edu.pl/temat/roznosci/biologia/2011/04/15/Ewolucja_geometryczna_czyli_jak_powstalo_oko/
(....) i tak dalej....

https://pl.wikipedia.org/wiki/Karol_Sabath
https://www.app.pan.pl/archive/published/app52/app52-828.pdf

„Karol Sabath (ur. 24 sierpnia 1963 w Katowicach, zm. 10 października 2007 w Warszawie[1]) – polski biolog, paleontolog, popularyzator nauki (zwłaszcza biologii ewolucyjnej i paleontologii), jeden z najbardziej aktywnych w Polsce krytyków kreacjonizmu i hipotezy inteligentnego projektu.”
W celu tego krytykowania i propagowania założył internetowy Serwis Ewolucyjny. Strona wisiała na serwerze dość długo po jego nagłej śmierci, pisywał tam też Paweł Poręba. W miarę postępów wiedzy ktoś usiłował ją modyfikować, uwspółcześniać, co okazało się wlewaniem młodego wina do starych bukłaków. Czas i postępy w wiedzy oraz bezlitosne ramię falsyfikacji w końcu zaorały zawarte tam argumenty i ktoś skasował bloga: http://www.ewolucja.org/
W internecie znalazłem jakiś klon Serwisu Ewolucyjnego, ale autorzy szybko stracili zapał – ocaliłem kilka artykułów z Serwisu Ewolucyjnego w plikach pdf, bo były mi potrzebne do moich artykułów: http://www.darwin.pan.pl/d2d.html
Np. ten, który Karol Sabath napisał w odwecie na moje zarzuty, ale po weryfikacji przez czas i nowe odkrycia okazały się zasadne ???? https://slawekp7.files.wordpress.com/2017/05/jednoc59bc487-ludzkiego-gatunku-nieuzasadniona-krytyka-_serwisu-ewolucyjnego_-mojego-wpisu-sprzed-11-lat-_.pdf
Argumenty z Serwisu Ewolucyjnego były takie, jak przekonania Richarda Dawkinsa na temat możliwości ewolucji oka. Kopiował z książek Richarda Dawkinsa w artykułach zamieszczanych w Wiedzy i Życiu, ale skasowali archiwum. Dziwny jest ten klon Serwisu ewolucyjnego. Podano na nim odnośnik do strony Karola Sabatha, ale jak się w niego kliknie, to wyskakuje ta strona:
http://www.ewolucja.org/
Mimo to na tej podróbie Serwisu Ewolucyjnego widnieje informacja, że autorami jest Katarzyna Adamala i Karol Sabath. PaniAdamala może tak, ale nie Sabath, ponieważ nie żyje:


Po śmierci Karola Sabatha Katarzyna Adamala wyjechała do USA i podjęła pracę badawczą u noblisty Jacka Szostaka. Badali możliwości abiogenezy, ale nie okryli się sławą. Wręcz odwrotnie:
https://frontlinegenomics.com/news/17272/nobel-laureates-make-mistakes-jack-szostak-retracks-nature-chemistry-paper/
'Nature' wycofuje pracę Jacka Szostaka dotyczącą abiogenezy z 2016 r.!
"Powtórne eksperymenty nie potwierdziły replikacji RNA bez udziału polimerazy. Cała hipoteza "najpierw replikacja" wzięła w łeb! Noblista czuje się zażenowany: "Byliśmy zaślepieni wiarą w nasze odkrycia, nie byliśmy rygorystyczni..." -twierdzi noblista!"
Być może kroiła się grubsza afera, ponieważ znane mi tego typu artykuły z komentarzami szybko poginęły z internetu. W archiwum Nature w dalszym ciągu można przeczytać nieszczęsny artykuł, w którego pisaniu obok Jacka Szostaka uczestniczyła pani Katarzyna Adamala:
https://www.nature.com/nchem/journal/v8/n10/abs/nchem.2551.html

Wikipedia nie odstaje:
https://pl.wikipedia.org/wiki/Nieredukowalna_z%C5%82o%C5%BCono%C5%9B%C4%87

Jeden z polskich filozofów bardzo merytorycznie skomentował argumentację Richarda Dawkinsa:
Jodkowski słusznie napisał o Richardzie Dawkinsie:
https://creationism.org.pl/artykuly/KJodkowski2


Jest i o ewolucji oka:






 :)
:)
:)

Richard Dawkins został okrzyknięty Rottweilerem Darwina, co miało go stawiać ponad możliwościami Thomasa Huxleja nazwanego Buldogiem Darwina: https://www.wielkiepytania.pl/article/thomas-huxley-buldog-darwina/
Określenia te były wynikiem tego, że zarówno Huxlej, jak i Dawkins bardzo się angażowali w obronę poglądów Karola Darwina. Richard Dawkins podczas tej obrony często posuwał się do granic absurdu, co widać powyżej i np. tutaj: https://doczz.pl/doc/568888/richard-dawkins-jako-etolog-ewolucyjny-%E2%80%9Ewyja%C5%9Bnia%E2%80%9D-na-jaki...
Dawkins nasadził tam takich kocopołów, że film z polskimi napisami nagle zniknął z youtuba:
https://youtu.be/r73Cu4IvOLE

Nie wszyscy biolodzy dali się zwieść. W tym wykładzie dla doktorantów biologii profesor Włodzimierz Korohoda stwierdził, że Richard Dawkins nie jest naukowcem, tylko dziennikarzem:
Poza tym Dawkins nie był taki oryginalny za jakiego wielu fanów go uznawało. Jego koncepcja Samolubnego genu, to ubrane w odpowiednie słowa osiągnięcia wybitnych biologów ewolucyjnych. Inni siali a on zżął laury :)
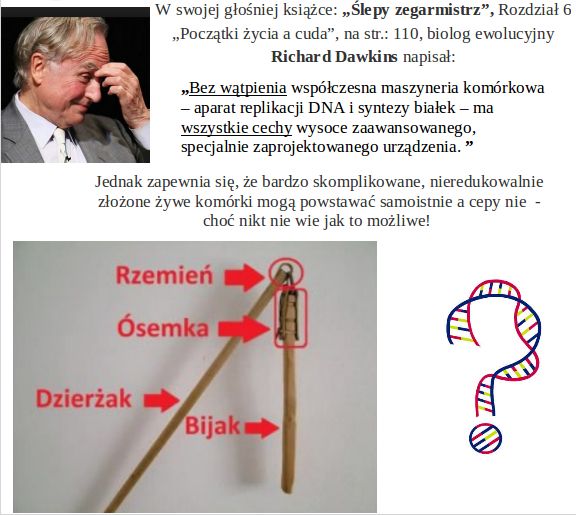
https://obserwatorlaicki.files.wordpress.com/2009/12/richard-dawkins-slepy-zegarmistrz-skan1.pdf

Niemniej Richard Dawkins potrafił też zaskakiwać. W tym przypadku chyba go poniosło, ponieważ w nowszych anglojęzycznych wydaniach "Ślepego zegarmistrza" nie ma już tego fragmentu:
Myśli Darwina przenikły do innych dziedzin niż nauki przyrodnicze

Darwinizm w przemyśle filmowym i muzycznym
Pomysły darwinistów nie skupiają się jedynie w obrębie rozważań w łonie biologii ewolucyjnej. Mają szeroki zasięg. Przeniknęły do filozofii, sztuki w tym do przemysłu filmowego. W taki sposób urabia się pogląd, że dla ewolucji nie ma rzeczy niemożliwych, że intuicje które skłaniają do buntu umysłowego wobec pomysłów darwinistów, to religijne złudzenie, a zaprzeczanie im nie opiera się na nauce tylko na osobistym niedowierzaniu, lub ignorancji! Film "Początek", to filozoficzna, bajkowa opowieść przyozdobiona w pseudonaukowy kitel:

https://www.filmweb.pl/reviews/recenzja-filmu-Pocz%C4%85tek-16160
"Biolog molekularny, u progu przełomu naukowego, podróżuje do Indii w poszukiwaniu dziewczyny, która może obalić lub potwierdzić jego teorię.
Amerykański reżyser Mike Cahill strzela na ekranie z takich armat, jakby kino miało się skończyć wraz z nim. Przeznaczenie kontra przypadek, religia kontra nauka, reinkarnacja kontra otchłań niebytu, miłość kontra społeczne posłannictwo, Richard Dawkins kontra Dalajlama. Wtłoczony w formułę niezależnego kina sci-fi"
Tutaj można obejrzeć "Początek":
https://www.cda.pl/video/180100ed
Darwinowsko-Dawkinkowska myśl przeniknęła też do przemysłu muzycznego:
https://www.newstatesman.com/culture/2015/02/richard-dawkins-feature-finnish-metal-band-nightwishs-new-album
Richard Dawkins pojawi się na nowym albumie fińskiego zespołu metalowego Nightwish!

ŹRÓDŁA
The master control gene for morphogenesis and evolution of the eye
https://pubmed.ncbi.nlm.nih.gov/9078363/
The genes involved in the morphogenesis of the eye
https://pubmed.ncbi.nlm.nih.gov/7905035/
Zwierzęce oczy
https://www.istockphoto.com/pl/obrazy/zwierz%C4%99ce-oko

https://pl.wikipedia.org/wiki/G%C5%82%C4%99boka_homologia
Z wyjaśnieniem będziemy mieli do czynienia, gdy nam darwiniści opowiedzą, na jakich zasadach ewoluowała "głęboka homologia" :)
The Eye of the Common Octopus (Octopus vulgari
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6971404/
Deep Homology?
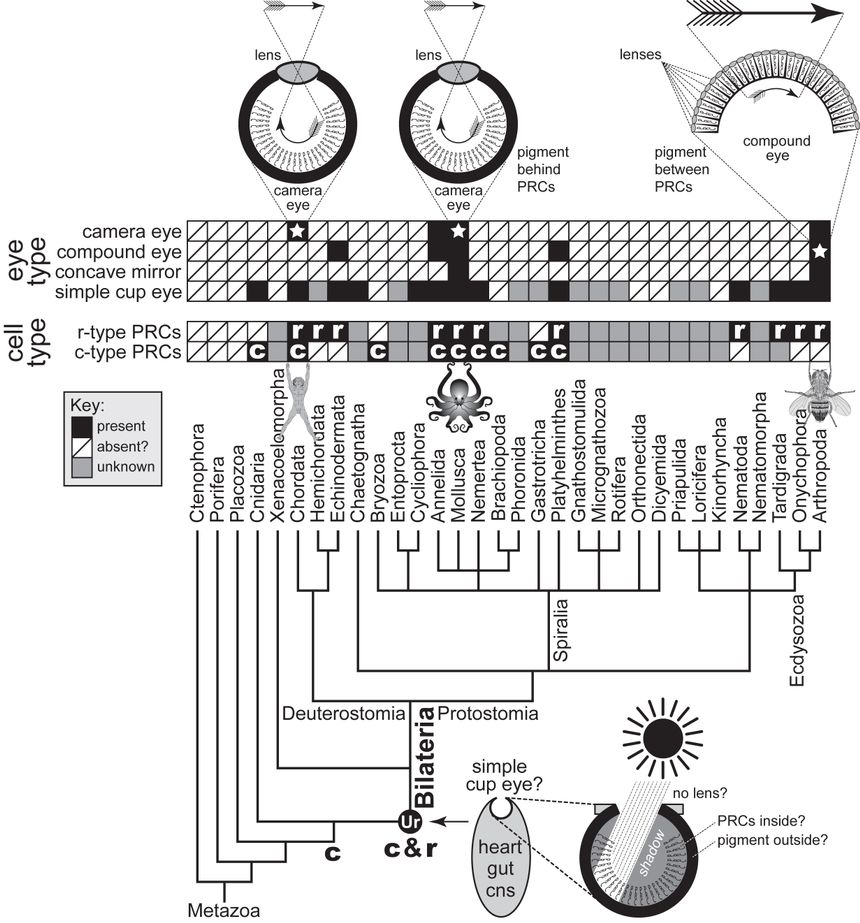
https://www.sdbonline.org/sites/fly/lewhelddeep/deephomology11.htm
Comparative Analysis of Gene Expression for Convergent Evolution of Camera Eye Between Octopus and Human
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC509264/
Comparative Analysis of Gene Expression for Convergent Evolution of Camera Eye Between Octopus and Human
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC509264/

Czym jest nieredukowalna złożoność w biologii - obszerne wyjaśnienie

Obszerne wyjaśnienie koncepcji nieredukowalnej złożoności systemów molekularnych
https://slawekp7.files.wordpress.com/2019/05/mechanizm-krzepnic499cia-krwi-e28093-system-nieredukowalnie-zc582oc5bcony-1-co-to-jest-nieredukowalna-zc582oc5bconoc59bc487-kompleksc3b3w-molekularnych-2-na-czym-polega-koncepcja-nieredu.pdf


http://cejsh.icm.edu.pl/cejsh/element/bwmeta1.element.desklight-cafb7118-f7e3-472b-a263-14878a2a2480/c/Behe__Nieredukowalna_zlozonosc....pdf