
https://pl.wikipedia.org/wiki/Mutacja_cicha
"Mutacja synonimiczna to zmiana pojedynczego nukleotydu w genie (mutacja punktowa) nie powodująca zmiany aminokwasu w kodowanym białku ze względu na to, że kod genetyczny jest zdegenerowany, czyli jeden aminokwas może być kodowany przez kilka kodonów (np. zmiana CCC na CCU nie powoduje zmiany aminokwasu, gdyż obie trójki kodują prolinę). Wydaje się, że w niektórych przypadkach, jeśli zmiana następuje z kodonu preferowanego na rzadki, może to wpłynąć na szybkość syntezy białka, a to z kolei na szybkość jego zwijania, a zatem na jego strukturę przestrzenną, co może mieć wpływ na fenotyp"

Dowody na adaptacyjne podstawienia synonimiczne, jako wyniki eksperymentalnych ewolucji drobnoustrojów
Susan F. Bailey, Luz Angela Alonso Morales , Rees Kassen Genome Biology and Evolution , tom 13,
wydanie 9, wrzesień 2021

„Często się zakłada , że mutacje synonimiczne są neutralne pod względem dostosowania do wymagań środowiska, ponieważ nie zmieniają kodowanego aminokwasu, a zatem nie mogą być „zauważone” przez dobór naturalny.
Jednak coraz więcej dowodów każe wnioskować, że mutacje synonimiczne mają wpływ na dostosowanie za pośrednictwem ewolucji adaptacyjnej, poprzez wpływ mutacji synonimicznych na ekspresję genów i fałdowanie białek.
W niniejszym artykule dokonujemy przeglądu tego, czego przeprowadzane przez licznych badaczy eksperymenty mikrobiologiczne nauczyły nas o roli i zaangażowaniu mutacji synonimicznych w mechanizmy adaptacyjne.
Przegląd opisanych w literaturze naukowej eksperymentów z mutagenezą ukierunkowaną ujawnił, że rozkłady efektów dostosowania dla mutacji niesynonimicznych i synonimicznych są bardziej do siebie podobne – szczególnie w przypadku mutacji korzystnych – niż się spodziewali biolodzy przekonani o neutralnym charakterze mutacji synonimicznych. Z tych korelacji wynika, że rola i zakres wkładu mutacji synonimicznych w możliwości adaptacyjne organizmów są częstsze niż by to wynikało z wcześniejszych obserwacji.
Przegląd prac opisujących wyniki badań nad mechanizmami ewolucji adaptacyjnej , pokazuje, że mutacje synonimiczne wpływają na dostosowanie poprzez szereg mechanizmów molekularnych:
- Tworzenie niespecyficznych miejsc wiązania się polimerazy RNA z wpływem na transkrypcję.
- Mutacje z wpływem na stabilność i strukturę [konformację] matrycowego RNA, co ma bezpośredni wpływ regulację ekspresji genów na poziomie translacji.
- Wpływ na konformacje i stabilność białek.
Mutacje synonimiczne nie zmieniają kodowanych aminokwasów, ale nadal mogą wpływać na kondycję organizmów poprzez wpływ na ekspresję genów i strukturę białek. Dowody pochodzą obecnie z szeregu badań obserwacyjnych, z dziedziny genomiki porównawczej i z wyników badań eksperymentalnych. Pojawia się coraz więcej dowodów na to, że niektóre mutacje synonimiczne mogą mieć duży wpływ na dostosowanie, a zatem mogą odgrywać znaczną rolę w ewolucji adaptacyjnej w znacznie krótszych skalach czasowych niż zakładają zwolennicy neodarwinowskiej wizji ewolucji [w wyniku powstawania losowych mutacji selekcjonowanych przez dobór naturalny].
W tej pracy przeglądowej analizujemy dowody pochodzące z dwóch rodzajów badań eksperymentalnych. Pierwsze to te, które bezpośrednio określają ilościowo przydatność zbioru mutantów synonimicznych i niesynonimicznych pod względem pozytywnej selekcji – zwykle generowanych przez ukierunkowaną mutagenezę – na każdym etapie ewolucji eksperymentalnej. Ponadto zajmiemy się opisami i analizą eksperymentatorów, którzy śledzą dynamikę ewolucyjną rozmnażanych populacji hodowanych – w zaprojektowanych, kontrolowanych środowiskach.
Wyniki tych eksperymentalnych badań ewolucji identyfikują mutacje synonimiczne i niesynonimiczne [i inne], które powstają podczas ewolucji eksperymentalnej. W tym celu naukowcy sekwencjonują i porównują całe genomy. Obserwują i opisują zakres dostosowania i stopień sprawności mutantów pod względem zdobywania przewagi selekcyjnej w wyniku mutacji. Omówione tutaj rodzaje badań zostały przeprowadzone na populacjach wirusów, bakterii lub drożdży, dlatego z konieczności koncentrujemy się na procesach adaptacyjnych obserwowanych u tych drobnoustrojów.
Zakończymy dyskusją na temat wpływu błędnego założenia, że mutacje synonimiczne są neutralne i omówimy kierunki przyszłych prac nad rolą mutacji synonimicznych w ewolucji adaptacyjnej.”
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3078729

"[….] Niedawno doniesiono, że synonimowe mutacje punktowe występują znacznie częściej niż niesynonimowe zmiany zasad w pseudogenie Drosophila Est-6 .
U kur zaobserwowano, że w obrębie wielu IglV i IghVpseudogenów, liczba kodonów stop zawartych w sekwencji „kodującej” jest znacznie niższa niż można by się spodziewać, gdyby podstawienia nukleotydów występowały losowo; ponadto większość kodonów stop wprowadzonych przez mutacje punktowe jest następnie „korygowana” i eliminowana przez kolejne mutacje punktowe w tym samym kodonie.
Ta cecha, która występuje również w V H pseudogenów u myszy, może wskazywać, że te pseudogeny są w stanie kodować białka, lub że ochrona otwartych ramek odczytu umożliwia pseudogenom zaangażowanie w rearanżacje genów somatycznych."
Dodam, że uczeni nie tylko zajmują się rolą mutacji synonimicznych w kontekście ich przydatności adaptacyjnych.Znaleziono związek między tym rodzajem mutacji a wieloma chorobami: kancerogenezą skutkującą rozwojem nowotworów i innymi:

„Przez długi czas synonimiczne polimorfizmy pojedynczego nukleotydu były uważane za nieme mutacje. Jednak obecnie dobrze wiadomo, że mogą one wpływać na konformację i funkcję białek, prowadząc do zmienionych podatności na choroby, zróżnicowanego rokowania i/lub odpowiedzi na lek.
Dzieje się to za pośrednictwem różnych mechanizmów:
zakłócając sygnały splicingu prekursorowych mRNA,
wpływając na miejsca wiązania regulatorów czynników transkrypcyjnych,
i miRNA,
lub modyfikując drugorzędową strukturę mRNA.
W tym artykule analizujemy 22 ludzkie choroby lub cechy genetyczne, powiązane z 35 synonimicznymi polimorfizmami pojedynczego nukleotydu w 27 różnych genach.
Przeprowadziliśmy analizę kontekstu sekwencji lokalnej pod kątem skłonności do pauz rybosomalnych, na które wpływają synonimiczne polimorfizmy pojedynczego nukleotydu.
Wnioski:
Wyniki te sugerują, że choroby genetyczne lub cechy związane z mutacjami synonimicznymi mogą być konsekwencją innego mechanizmu leżącego u podstaw nie-synonimicznych mutacji.”
https://pubmed.ncbi.nlm.nih.gov/33836673/
Mutacje synonimiczne regulujące szybkość translacji mogą odgrywać istotną rolę w rozwoju raka wątroby
Qun Li 1, Jian Li 1, Chun-Peng Yu 1, Shuai Chang 1, Ling-Ling Xie 1, Piosenka Wang 2
https://pubmed.ncbi.nlm.nih.gov/24630730/
https://pubmed.ncbi.nlm.nih.gov/26300548/
Funkcjonalne znaczenie somatycznych mutacji synonimicznych w czerniaku i innych nowotworach
Valer Gotei 1, Jared J Gartner 2, Nouar Qutob 3, Laura Elnicki 1, Yardena Samuels
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5085179/
Synonimiczne mutacje w onkogenezie i apoptozie
Xiang Li , 1 Yuan Chen , 1 Hong Qi , 2 Liyu Liu , 3 i Jianwei Shuai 1, 4
https://pubmed.ncbi.nlm.nih.gov/21878961/
Wkład mutacji synonimicznych w choroby u ludzi
Sauna Zuben E 1, Chava Kimchi-Sarfaty
Niektórzy badacze, w tym przypadku bioinformatyk, muszą nauczyć się czekać :)
https://pl.sci.biologia.narkive.com/FFf0NisO/pytanie-do-nie-ewolucjonistow#post2
"Przemol", to stary weteran grupy pl.sci.biologia i obrońca darwinizmu. Po dziś dzień pisuje na tą grupę stojąc na posterunku misji, jaką obrał! :)




Istnieje alternatywne wyjaśnienie panie Weiner:
Tego pana najbardziej poniosło :)
"Paweł Jałoszyński – polski biotechnolog i entomolog specjalizujący się w koleopterologii." [za wikipedią]

Kupę lat temu w rozmowach z biologami ewolucyjnymi wałkowałem temat ewolucji przeobrażenia zupełnego. Między innymi rozmawiałem z Karolem Sabathem - założycielem 'Serwisu Ewolucyjnego' Pan Karol wskazywał na korzyści, twierdził że nie czyta literatury specjalistów od rozwoju post-embrionalnego bezkręgowców, aż w końcu przyznał, że nie ma teorii wyjaśniającej przebieg ewolucji holometabolii i że anty-darwiniści, którzy domagają się tego typu modeli teoretycznych są ordynarni i stosują wytrychy retoryczne. Za jakiś czas zlecił panu Pawłowi Jałoszyńskiemu napisanie czegoś na ten temat, a jak napisał zostałem o tym fakcie poinformowany. Pan Paweł streścił oklepaną bajkę - nie ma już 'Serwisu Ewolucyjnego', ale opowieść pana Pawła Jałoszyńskiego została uratowana!
https://slawekp7.files.wordpress.com/2012/04/ewolucja-metamorfozy-u-owadc3b3w.pdf
Jak łatwo można się przekonać odpowiedź pana Jołoszyńskiego nie odnosi się do moich pytań, a tym bardziej nie wyjaśnia, jak wyewoluowało przeobrażenie zupełne u owadów:
https://slawekp7.wordpress.com/2021/08/21/od-gasienicy-do-motyla-cudowny-proces-holometabolii/
W tej dyskusji udzielił się Kuba - słynny quasi-biolog. Niestety już nie żyje - mimo radykalnie odmiennych poglądów Kuba dał się lubić:
https://www.wykop.pl/wpis/6831784/legendarny-post-quasiego-z-salonu24-przytaczam-gdy/
https://msp.money.pl/grupa-pl_soc_prawo/samobojstwo;-;nie;rozumiem,watek,215868.html

[....]
Kuba w tym świecie już się nie dowie, że człowiek z którego szydzi miał rację! quasi (biolog) uznał jego komentarz za ewidentny7 przykład kreacjonistycznego, błędnego myślenia - było to myślenie naukowe :)


https://www.znaczacy.com/pytanie-do-nie-ewolucjonistow-t848990/2

ZAISTE!

Coś takiego panie January - na końcu niniejszego tekstu jest o tym, jak sobie radzi! :)

Szkoda, że Kuba nie dożył do naszych czasów :(
https://slawekp7.wordpress.com/2021/02/12/gggggggo/

[.....]

[.....]

Enzo Salviati - miło wspominam tego biologa (edytora wikipedii). Czasami mnie drażnił, bo potrafił być uszczypliwy. Niemniej rekompensował oryginalnym poczuciem humoru. Poniżej napisał całkiem poważnie! :)
https://www.znaczacy.com/pytanie-do-nie-ewolucjonistow-t848990/3


Należy przyznać, że Enzo dostarczył wyjaśnienia pochodzenia nieredukowalnej złożoności w biologii! Hipoteza pod tytułem:


Oczywiście nie jest to wyjaśnienie oryginalne :)
https://slawekp7.wordpress.com/2021/08/18/ktore-obrazki-pomagaja-lepiej-zrozumiec-argumentacje-allena-orra-i-michaela-behego/



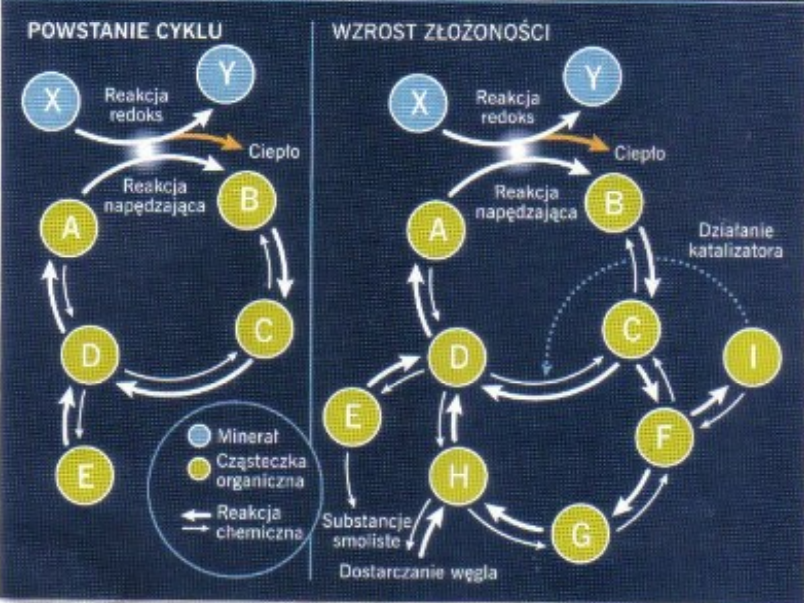
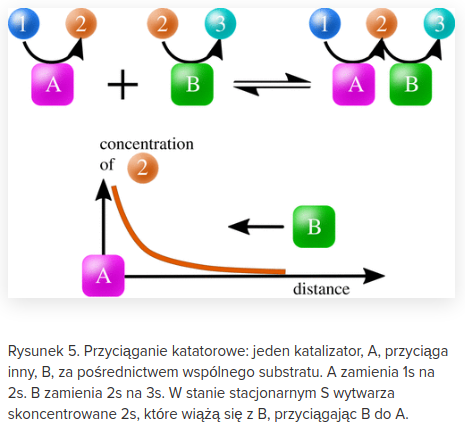
Jest to wyjaśnienie, do którego Michael Behe się odniósł, o czym Enzo Salviati został poinformowany na długo przed tą dyskusją. Zanim się rozwinęła Enzo dłuższy czas dyskutował ze mną na privie. Poza tym Enzo mi powiedział, że widział Michaela Behe'ego na jakimś sympozjum poświęconym biologii ewolucyjnej, zatem miał okazję dopytać go osobiście :)



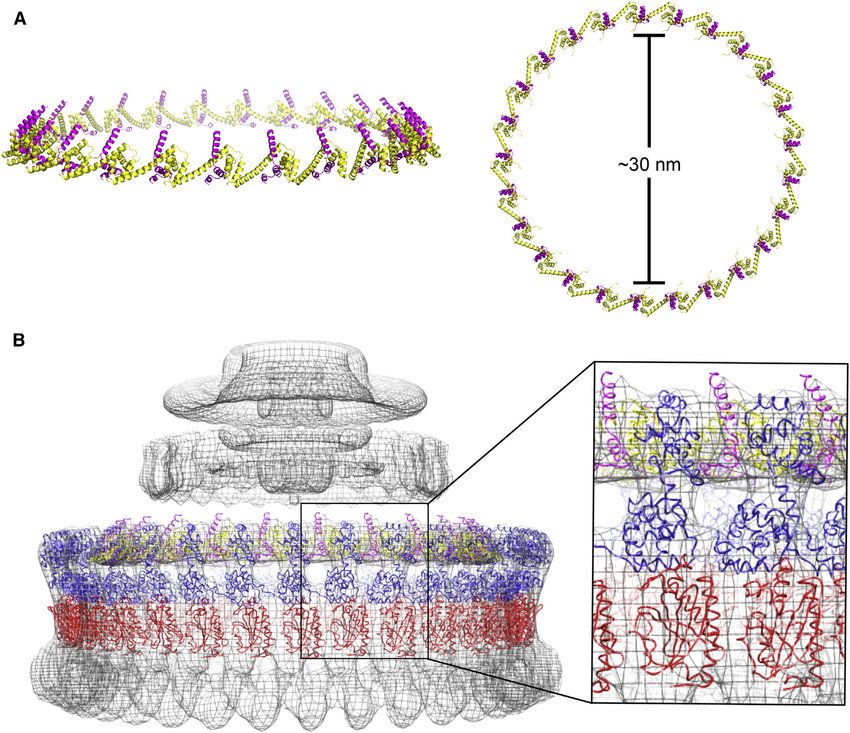
PODSUMOWANIE:

Nic dodać, nic ująć :)

https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3078729
„[….]
Niedawno doniesiono, że synonimowe mutacje punktowe występują znacznie częściej niż niesynonimowe zmiany zasad w pseudogenie Drozofila Est-6 .
U kur zaobserwowano, że w obrębie wielu IglV i IghVpseudogenów, liczba kodonów stop zawartych w sekwencji „kodującej” jest znacznie niższa niż można by się spodziewać, gdyby podstawienia nukleotydów występowały losowo; ponadto większość kodonów stop wprowadzonych przez mutacje punktowe jest następnie „korygowana” i eliminowana przez kolejne mutacje punktowe w tym samym kodonie.
Ta cecha, która występuje również w V H pseudogenów u myszy, może wskazywać, że te pseudogeny są w stanie kodować białka, lub że ochrona otwartych ramek odczytu umożliwia pseudogenom zaangażowanie w rearanżacje genów somatycznych.”
Pseudogeny: Pseudo-funkcjonalne czy kluczowe regulatory w zdrowiu i chorobie?

„Pseudogeny od dawna są określane jako „śmieciowe” DNA, nieudane kopie genów, które powstają podczas ewolucji genomów [lub takie, które po awarii wyłączają całe sieci genetyczne – np. te, które rzekomo przyczyniły się do utraty zębów u ptaków czy mrówkojadów].
Jednak ostatnie wyniki badań podważają ten pseudogenizm; w rzeczywistości niektóre pseudogeny wydają się mieć potencjał do regulowania swoich kodujących białka kuzynów [genów]. Dalekie są od bycia cichymi reliktami, wiele pseudogenów ulega transkrypcji do mRNA, niektóre wykazują specyficzny wzór aktywacji w tkankach.
Transkrypty pseudogenów mogą być porzetwarzane w krótkie, interferujące RNA, które regulują ekspresję kodujących genów, poprzez szlak RNAi.
W innym niezwykłym odkryciu wykazano, że pseudogeny są zdolne do regulowania supresorów nowotworowych i onkogenów, działając jako wabiki mikroRNA. Odkrycie, że pseudogeny często ulegają deregulacji podczas progresji nowotworu, gwarantuje dalsze badanie prawdziwego zakresu funkcji pseudogenów.
Genom ludzki, podobnie jak u innych ssaków, jest zaśmiecony różnorodnymi powtarzającymi się elementami i niekodującymi genami. Jednym z takich elementów jest pseudogen, zaburzone mutacjami punktowymi i pocięte delecjami odwzorowanie oryginalnego genu kodującego białko, który utracił zdolność do wytwarzania funkcjonalnego białka. Ponieważ nie kodują białek, często zakłada się, że pseudogeny są niefunkcjonalne i zaliczane do „śmieciowego DNA”.
Podczas gdy niektóre pseudogeny są ciche transkrypcyjnie, inne są aktywne, co rodzi pytanie, czy ich niekodujące transkrypty bezsensownie zużywają energię komórki, czy zamiast tego są wykorzystywane przez komórkę do regulacji kodujących genów. To pytanie jest szczególnie istotne, biorąc pod uwagę niedawną lawinę dowodów sugerujących, że długie niekodujące mRNA odgrywają kluczową rolę w regulacji funkcji genomu.Pseudogeny mogą powstawać poprzez różne mechanizmy. Spontaniczne mutacje w genie kodującym, które uniemożliwiają transkrypcję lub translację genu – prowadzą do powstania „jednolitego” pseudogenu . Zduplikowane pseudogeny są tworzone przez duplikację tandemową, lub nierówny crossing-over. Te zduplikowane geny tracą swój potencjał kodowania białek z powodu utraty promotorów, lub wzmacniaczy, albo okaleczających je mutacji, takich jak przesunięcia ramki odczytu, lub powstawanie przedwczesnych kodonów stop.
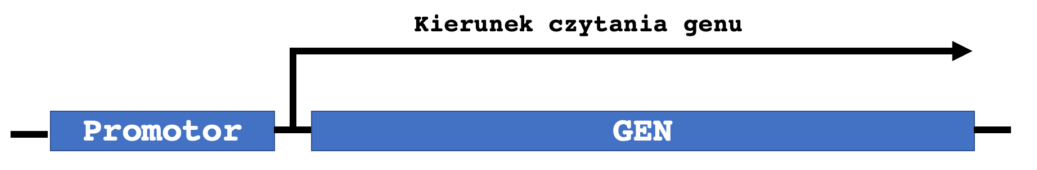
Jednak mają tendencję do zachowywania swojej charakterystycznej struktury intron-egzon. W przeciwieństwie do tego, retrotransponowane, lub „przetworzone” pseudogeny (PP) są wytwarzane, gdy transkrypt mRNA jest poddawany odwrotnej transkrypcji i integrowany z genomem w nowej lokalizacji, a zatem w tym przypadku nie zawierają intronów.Inne wspólne cechy PP to ich ciągi poliA i bezpośrednie powtórzenia na każdym końcu pseudogenu. Wydaje się, że w retrotranspozycji mRNA do DNA i integracji z genomem pośredniczy długi, rozproszony element jądrowy 1 (L1), a aktywność transkrypcyjna powstałego PP zależy od tego, czy zdarzenie integracji zachodzi blisko innego promotora. Zbiór przetworzonych pseudogenów w ludzkim genomie został wygenerowany z zaledwie 10% genów kodujących. Wysoce eksprymowane geny porządkowe z większym prawdopodobieństwem produkują PP, podobnie jak inne krótsze RNA o wysokim stopniu transkrypcji. Przykładem jest niewielka liczba genów kodujących białka rybosomalne, które stanowią około 20% ludzkich PP .Termin pseudogen został ukuty w 1977, kiedy Jacq i współpracownicy odkryli wersję genu kodującego 5S rRNA, który został skrócony, ale zachował homologię z aktywnym genem Xenopus laevis. W ciągu następnych dwóch dekad pseudogeny odkryto sporadycznie. Przyspieszenie technologii sekwencjonowania nowej generacji w połączeniu z projektem senkwencjonowania ludzkiego genomu pozwoliło uzyskać obrazy pełnego genomu szeregu organizmów, umożliwiając znacznie dokładniejsze analizy częstości występowania pseudogenów.
Co godne uwagi, pseudogeny są prawie tak liczne jak geny kodujące, a prognozy wahają się od 10 000 do 20 000 ludzkich pseudogenów. Większość ludzkich pseudogenów to PP, podczas gdy liczba jednostkowych pseudogenów w ludzkim genomie wynosi <100.Pseudogeny są obecne w wielu gatunkach, w tym roślinach, bakteriach – choć nie są tak liczne w organizmach jednokomórkowych w owadach i nicieni , ale są one szczególnie liczne u ssaków. Pseudogeny są czasami uważane za reprezentujące „sekwencję neutralną”, w której kumulujące się mutacje nie są selekcjonowane szkodząc, lub wspierając organizm – skoro miały nie mieć wpływu na fenotyp, to tym samym z punktu widzenia darwinizmu dobór naturalny ich nie zauważa. Jednak ta przesłanka opiera się na założeniu, że pseudogeny są funkcjonalnie obojętne. Ostatnio pojawiły się dowody na to, że niektóre pseudogeny są funkcjonalnie aktywne, a zatem badanie ich ewolucji i przyczyn konserwowania przez naturalną selekcję może wesprzeć przekonanie o ich użyteczności dla organizmu dać wgląd w ich potencjalny mechanizm działania.
Niedawno doniesiono, że synonimowe mutacje punktowe występują znacznie częściej niż niesynonimowe zmiany zasad w pseudogenie Drosophila Est-6 .
U kur zaobserwowano, że w obrębie wielu IglV i IghVpseudogenów, liczba kodonów stop zawartych w sekwencji „kodującej” jest znacznie niższa niż można by się spodziewać, gdyby podstawienia nukleotydów występowały losowo; ponadto większość kodonów stop wprowadzonych przez mutacje punktowe jest następnie „korygowana” i eliminowana przez kolejne mutacje punktowe w tym samym kodonie.
Ta cecha, która występuje również w V H pseudogenów u myszy, może wskazywać, że te pseudogeny są w stanie kodować białka, lub że ochrona otwartych ramek odczytu umożliwia pseudogenom zaangażowanie w rearanżacje genów somatycznych.
Pseudogeny stopniowo gromadzą mutacje, a liczba mutacji może dać nam oszacowanie ich wieku. (….) Analiza regionu rozszerzonego układu zgodności tkankowej makaka (MHC) klasy II ujawniła dwa pseudogeny, które okazały się homologiczne do ludzkich pseudogenów HIV typu TAT-podobnych do czynnika 1 i palca cynkowego, co sugerowało konserwację ewolucyjną tych sekwencji.
Badania prowadzone przez Podlaha i współpracowników wykazały, że pseudogen Makorin1-p1 jest zachowany w szczepach Mus musculus i Mus pahari . To skłoniło specjalistów do badania całych genomów i poszukiwania pseudogenów zachowanych między ludźmi a myszami. Ludzkie pseudogeny, wraz z ich genami macierzystymi, zostały porównane z odpowiadającymi im ortologami myszy i ich pseudogenami.
Co ciekawe, stwierdzono, że wiele z badanych pseudogenów nagromadziło bardzo niewiele mutacji w regionach regulatorowych w porównaniu z genami macierzystymi, co może sugerować, że te regiony regulatorowe mają znaczenie dla pseudogenu i że pseudogen może być funkcjonalny.
Spośród badanych grup genów i pseudogenów analiza sekwencji sugerowała, że 30 z nich reprezentuje pseudogeny, które były obecne zarówno u myszy, jak i u ludzi i powstały przed rozdzieleniem się tych dwóch gatunków. Porównanie transkrybowanych ludzkich pseudogenów pokazuje, że 50% jest konserwatywnych u rezusów, ale tylko 3% jest konserwatywnych u myszy.
Analizy tych pseudogenów wykazały, że pomimo K A / K S szybkość substytucji wskazująca na niekodujące RNA, poziomy GC i szybkość mutacji w tych pseudogenach jest ograniczona w stosunku do otaczających je regionów międzygenowych. Są to liczne dowody na to, że niektóre pseudogeny wykazują funkcjonalną rolę w organizmach, które je przechowują.
Większość pseudogenów traci zdolność do transkrypcji, albo z powodu mutacji w ich promotorze, albo (w przypadku PP) integracji z cichymi regionami genomu. Dokonywanie dokładnych pomiarów transkrypcji pseudogenów jest skomplikowane ze względu na podobieństwo, jakie dzielą one z genami macierzystymi .
Jednakże istnieje wiele przykładów pseudogenów, które podlegają transkrypcji, w tym pseudogeny dla supresora nowotworu PTEN (którego transkrypty są liczniejsze niż gen macierzysty), nadnerczowej hydroksylazy sterydowej P450c21A, ludzki interferon leukocytów, GAPDH , glukocerebrozydaza i Oct4. Technologia mikromacierzy umożliwia analizę transkrypcji pseudogenów na znacznie większą skalę. Szacunki dotyczące proporcji transkrybowanych ludzkich pseudogenów wahają się od 2% do 20%.
Badanie wzorca transkrypcji w tkankach i liniach komórkowych może dać wgląd w potencjalną funkcjonalność. Inne niekodujące RNA wykazują specyficzne dla tkanki wzorce ekspresji i wykazano również, że pełnią role funkcjonalne, w tym antysensowne RNA , transkrypty międzygenowe i długie niekodujące RNA oraz miRNA .
W badaniu transkrypcji w regionach ENCODE genomu stwierdzono transkrypcję 14 pseudogenów. Pięć z nich zostało przepisanych wyłącznie w jądrach, a kolejne cztery były również aktywne w jądrach i innych tkankach. Ten wzór transkrypcji jest zgodny z wcześniejszymi wynikami, co sugeruje możliwe znaczenie biologiczne dla specyficznej dla jąder transkrypcji pseudogenów. Istnieje również kilka przykładów pseudogenów, których czasowo-przestrzenny wzorzec ekspresji różni się od wzorca ich genu macierzystego.
Specyficzne zmiany w ekspresji pseudogenów mogą również wystąpić w różnych warunkach fizjologicznych, w tym w chorobach takich jak cukrzyca i rak. Przykłady dynamicznej transkrypcji pseudogenów zaobserwowano w innych organizmach. Silną transkrypcję pseudogenów zmierzono w Mycobacterium leprae, organizmie wywołującym trąd, przy poziomach specyficznych transkryptów zmieniających się podczas procesu infekcji. Indukcja stresu u Arabidopsis thaliana prowadzi do zmian w ekspresji wielu genów i pseudogenów.
Aktywność transkrypcyjna pseudogenu będzie częściowo zależeć od wykorzystywanego przez niego promotora. Niektóre mają własne promotory, podczas gdy inne wykorzystują promotory pobliskich genów. Przetworzony pseudogen jest w dużej mierze na łasce swojego miejsca integracji, jeśli chodzi o aktywność promotora. Dlatego też różnica we wzorcu transkrypcji między pseudogenem a jego genem macierzystym niekoniecznie odzwierciedla rolę funkcjonalną, ale zamiast tego może być jedynie wynikiem kierowania przez nowy promotor.
Gdyby to ostatnie było prawdą, można by przewidzieć, że akt transkrypcji pseudogenu byłby ewolucyjnie neutralny (lub nawet wybrany przeciwko oszczędzaniu energii komórkowej). Analiza konserwatywnych pseudogenów transkrybowanych pokazuje, że około 50% jest rzeczywiście zachowanych przez miliony lat ewolucji naczelnych (chociaż znacznie mniej jest zachowanych między gatunkami bardziej odległymi od człowieka, takimi jak gryzonie).
Fakt, że transkrypcja niektórych pseudogenów jest specyficzna tkankowo, dynamiczna i utrzymywana od tysiącleci sugeruje, że ich transkrypty mogą odgrywać pewną funkcjonalną rolę w komórkach. Nic nie zastąpi jednak eksperymentów funkcjonalnych, aby sprawdzić, czy pseudogeny i ich transkrypty mają bezpośrednią aktywną rolę.
Wydaje się, że wiele organizmów wielokomórkowych zachowuje obecność pseudogenów i nakłada ograniczenia na same sekwencje – uniemożliwia ich ekspresję. Z drugiej strony, różne organizmy jednokomórkowe aktywnie wydalają geny, które uległy pseudogenizacji. Pozostaje zatem pytanie, jaka jest potencjalna korzyść dla ssaków i innych złożonych organizmów z zatrzymywania i prawdopodobnie obejmowania ochroną genów, które utraciły potencjał kodowania białek?
Jedną z proponowanych funkcji pseudogenów jest działanie jako źródło różnorodności genetycznej, na przykład w wytwarzaniu przeciwciał lub zmienności antygenu. Jednak kontekście niekodującego RNA pseudogeny oferują potencjalnie znacznie bardziej dynamiczne mechanizmy regulacji dynamicznych procesów jądrowych. W ostatniej dekadzie ujawniono nowy poziom złożoności w regulacji ekspresji genów i funkcji jądra. Wykazano, że wiele niekodujących RNA, zarówno długich, jak i krótkich, reguluje różne procesy w komórkach. Wydaje się, że niekodujące RNA wytwarzane z niektórych pseudogenów są wykorzystywane przez różne fascynujące mechanizmy do kontrolowania funkcji genów.
(….)

Polecane:
W dwóch przełomowych doniesieniach wykazano, że w mysich oocytach części wielu transkryptów pseudogenów są przetwarzane na małe interferujące RNA (siRNA). Te siRNA pochodziły z pseudogenów o wewnętrznej strukturze drugorzędowej lub z dsRNA składających się z sensownych i antysensownych transkryptów (zaobserwowano kombinacje mRNA pseudogen-pseudogen i kodujący pseudogen). Utrata Dicer (białka niezbędnego do produkcji siRNA) doprowadziła do spadku poziomów siRNA pochodzących z pseudogenów i wzrostu poziomu kodowania mRNA genów z homologią z sekwencjami siRNA.
Sugeruje to, że siRNA wytworzone z dsRNA są zdolne do tłumienia ekspresji genów. Na przykład siRNA wygenerowane ze struktury spinki do włosów w pseudogenowym RNA Au76 były w stanie zahamować ekspresję z rodzicielskiego genu kodującego Rangap1 ( Watanabe i wsp. 2008). Wykryto kilka siRNA niosących podobieństwo sekwencji do kompleksu deacetylazy histonowej, HDAC1.
Wszystkie sekwencje siRNA pochodziły z serii pseudogenów HDAC1 (brak z samego genu HDAC1 ), jednak po nokaucie Dicer poziomy mRNA HDAC1 wzrosły, co sugeruje, że gen kodujący jest regulowany przez kompleks wyciszający indukowany przez RNA (RISC). ). W innym przypadku siRNA wytworzono z regionów dsRNA utworzonych między mRNA Ppp4r1 a antysensownym RNA wytworzonym z pseudogenu o 90% homologii.
siRNA wytworzone z tego parowania wydają się tłumić Ppp4r1gen. Podobne badanie na gatunku ryżu pokazuje, że niewielka część pseudogenów ulega transkrypcji i przetwarzaniu w siRNA po sparowaniu z genem kodującym lub paralogicznym transkryptem pseudogenu. Odkrycia te sugerują potencjalny mechanizm działania transkryptów antysensownych. Jednak okaże się, czy podobne procesy zachodzą w komórkach somatycznych ssaków.
(…..)
miRNA to klasa niekodujących RNA, które wpływają na stabilność mRNA. W ich specyficzności i funkcji pośredniczy parowanie zasad z celem (głównie w 3′ UTR); ich głównym efektem jest powodowanie degradacji mRNA, a tym samym obniżanie poziomów ekspresji. W niedawnym raporcie wykazano, że para gen-pseudogen jest koregulowana przez te same miRNA.
PTEN jest supresorem guza, który często ulega mutacji w jednym allelu w momencie pojawienia się raka. Podatność na raka zależy od dawki PTEN. Dlatego utrzymanie precyzyjnych poziomów białka PTEN ma kluczowe znaczenie dla zapobiegania onkogenezie. PTENP1 , pseudogen supresora nowotworu PTEN , podlega transkrypcji na wysokim poziomie.
(……)
Sugeruje to, że pseudogen PTENP1 działa jako „wabik miRNA”, wiążąc się i tym samym zmniejszając efektywne stężenie komórkowe miRNA, umożliwiając tym samym ucieczkę PTEN z represji, w której pośredniczy miRNA. Funkcjonalne powiązanie między parą PTEN / PTENP1 jest zgodne z odkryciami, że ich poziomy są często skorelowane w próbkach raka prostaty i że delecje ogniskowe zawierające PTENP1 występują często w sporadycznych przypadkach raka okrężnicy.
Podobny związek wykazano również między onkogenem KRAS a jego pseudogenemKRASP1. Co ciekawe, zdolność pseudogenu HMGA1-p do destabilizacji mRNA HMGA1 zależy od regionu 3′ UTR, co sugeruje, że zarówno pozytywne, jak i negatywne czynniki stabilizacji są zdolne do konkurencyjnej interakcji z nieulegającymi translacji regionami genów i pseudogenów do regulowania wydajności ekspresji.
(…..)
W dużej mierze pseudogeny zostały przeoczone w dążeniu do zrozumienia biologii zdrowia i choroby do tego stopnia, że sondy pseudogenów są często nieobecne w dostępnych na rynku mikromacierzach. Ponieważ pojawiają się dowody, że pseudogeny ulegają deregulacji w chorobie i że ich deregulacja może przyczyniać się do chorób takich jak cukrzyca i rak, powszechne nastawienie, że są to niefunkcjonalne relikty, powoli się zmienia.
Wraz z pojawieniem się niedrogiego sekwencjonowania nowej generacji, badania transkryptomiki, a w szczególności pseudogenów (i innych transkrybowanych elementów niekodujących), powinny doświadczyć kwantowego skoku naprzód. W nadchodzącym dziesięcioleciu zakres i mechanizmy funkcji pseudogenów powinny stać się jaśniejsze.”
ŹRÓDŁO
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3078729
Konwergencja molekularna
https://pl.sci.biologia.narkive.com/gCb5kk5d/podobienstwa-nie-musza-swiadczyc-o-pokrewienstwach
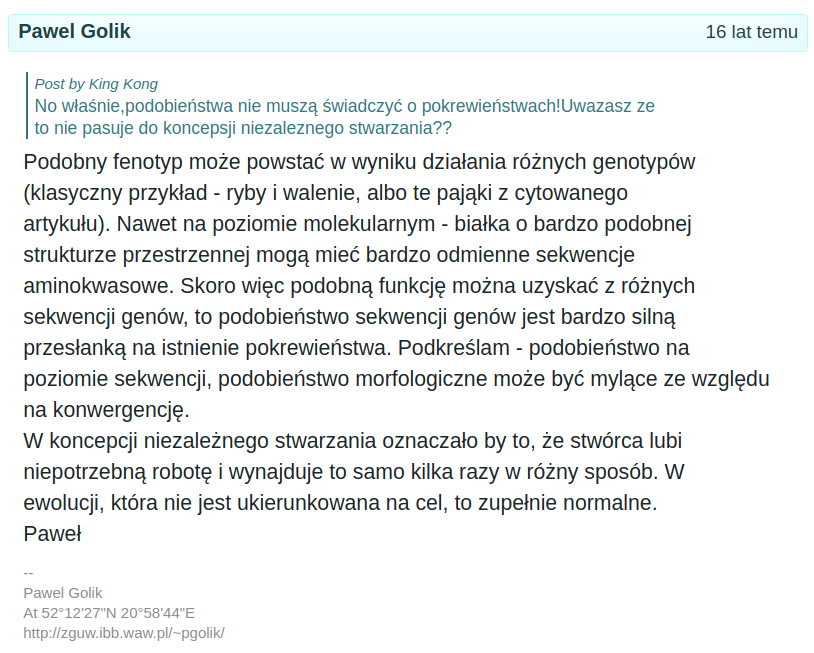
Np.:
https://www.newscientist.com/article/dn26694-humans-and-birds-share-the-same-singing-genes/
„Nowo zbadane genomy zostały wykorzystane do skonstruowania ostatecznego drzewa ewolucyjnego ptaków oraz do zbadania innych aspektów ewolucji ptaków – w tym tego, jak i kiedy ptaki straciły zęby.
Ale chociaż wysiłki konsorcjum oznaczają, że wiemy o wiele więcej o biologii ptaków niż w zeszłym tygodniu, wciąż pozostają pytania, na które należy odpowiedzieć.
Na przykład nie jest jasne, dlaczego istnieje tak niezwykły poziom podobieństwa genetycznego i na poziomie anatomii mózgów między ptakami uczącymi się śpiewu i ludźmi uczącymi się mowy, biorąc pod uwagę, że ostatni wspólny przodek tych dwóch grup mógł żyć 310 milionów lat temu. Czy konwergencja genetyczna jest zbiegiem okoliczności, czy też nauka głosu jest możliwa tylko wtedy, gdy obejmuje te 55 genów?”
https://science.sciencemag.org/content/346/6215/1256846
[…..] w kanale sodowym dla organów elektrycznych w niezależnych rodach ryb ( 29); oraz w genie prestyny do echolokacji u nietoperzy i waleni ( 30 ). Ponadto, w przypadku ssaków posługujących się echolokacją, zbieżność sekwencji aminokwasów w skali genomu została niedawno odkryta w wielu genach zaangażowanych w słyszenie ( 31 ). Podobnie, ostatnio donoszono o zbieżnych zmianach sekwencji aminokwasów u ptaków i ssaków uczących się głosu ( 32 , 33 ). Nie znamy jednak przykładów szerokiej zbieżności wzorców ekspresji genów w obszarach mózgu zaangażowanych w zbieżnie wyewoluowane cechy behawioralne.
Zbadaliśmy, czy złożona konwergencja behawioralna i anatomiczna jest związana z konwergencją molekularną u uczących się śpiewu ptaków oddalonych od wspólnego przodka o ponad 68 milionów lat i ludzi i ptaków – gatunków oddalonych ewolucyjnie o 310 milionów lat.
Nie chodzi jedynie o analogiczne wzory ekspresji genów i fenotypy na poziomie mózgów!
https://science.sciencemag.org/content/346/6215/1256846
Doszliśmy do wniosku, że jeden lub więcej genów leżących u podstaw złożonej cechy może wykazywać zbieżną ewolucję między gatunkami, nawet tymi oddalonymi o dziesiątki do setek milionów lat od wspólnego przodka. Na przykład, zbieżne identyczne podstawienia aminokwasów zostały zidentyfikowane w genie opsyny dla absorpcji światła w różnych liniach kręgowców ( 25 ); w receptorze melanokortykotropiny u ssaków i ptaków ( 26 ); w genie kodującym żółte plamki na skrzydłach używanych do zalotów u much ( 27 ); w czynnikach transkrypcyjnych Pitx w celu uzyskania i utraty kończyn u ryb i ssaków morskich ( 28 ); […..]
Identyczne krótkie sekwencje peptydowe w niepowiązanych białkach mogą mieć różne konformacje: poligon testowy dla teorii rozpoznawania immunologicznego
Ewolucja zbieżna w elementach strukturalnych białek badanych za pomocą analizy profili krzyżowych
https://pubmed.ncbi.nlm.nih.gov/22244085/
Białka o tej samej fałdzie i niepowiązanych sekwencjach mają podobny skład aminokwasowy
https://pubmed.ncbi.nlm.nih.gov/16565950/
ttps://royalsocietypublishing.org/doi/10.1098/rstb.2018.0234
U owadów, O konwergentnej ewolucji genetycznej niespokrewnionych grup owadów:
http://genomics-pubs.princeton.edu/insect_genomics/home.shtml

„Daleko od przypadkowości, ewolucja przebiega według przewidywalnego wzorca genetycznego – stwierdzili naukowcy z Princeton!”
W skład mechanizmów odpowiedzialnych za powstawanie polimorfizmu w populacji wchodzi szeroki wachlarz zjawisk należących do zbioru NORM REAKCJI NA SRODOWISKO.
Należy wymienić mechanizmy, które pokazał Grzegorz Mendel (zmiana frekwencji genów w wyniku dryfu genetycznego), powstawanie alleli wielokrotnych w wyniku mutacji i rekombinacji genetycznej (są to jedynie różne warianty tych samych genów)
Coraz lepiej poznawane mechanizmy epigenetyczne, czy oparte na nielosowych mutacjach (w tzw. gorących miejscach mutacyjnych) i selekcji oraz na zasadzie powtarzalnej zmienności (badania podczas hodowli mutacyjnych wykazały, że w kolejnych pokoleniach powstaja te same mutanty) – na podstawie tego mechanizmu niespokrewnione gatunki na naszych oczach nabierają odporności na te same toksyny, za pośrednictwem mutacji w analogicznych genach!
„Daleko od przypadkowości, ewolucja przebiega według przewidywalnego wzorca genetycznego – stwierdzili naukowcy z Princeton!
Naukowcy z Princeton University pod kierunkiem Petera Andolfatto, profesora nadzwyczajnego ekologii i biologii ewolucyjnej oraz Lewis-Sigler z Institute for Integrative Genomics odkryli, że ewolucją może kierować proste i powtarzalne rozwiązanie genetyczne wymuszone presja srodowiska, o szerokim zasiegu geogragicznym, ktora u niespokrewnionych gatunkow wywoluje takie same.zmiany genetyczne.
Oznacza to, że naukowcy posiadający wiedzę o tym, jak pewne warunki zewnętrzne wpływają na geny kodujace okreslone białka u różnych gatunków mogą zidentyfikowac mechanizmy genetyczne odpowiedzialne za adaptacje.
Wiele roznych gatunkow owadów niezależnie rozwinęło zdolność odżywiania się roślinami, które wytwarzają toksyny zwane kardenolidami i mogą je wykorzystywac do obrony przed drapieżnikami.
Badalismy docelowe bialko dla kardenolidów: Na +, K + -ATPazy u 14 gatunków owadow nalezacych do trzech rzedow, które żywią się roślinami produkującymi ta toksyne, podzielonych na 15 odseparowanych grup.
Pomimo dużej liczby potencjalnych celów modulowania stopnia wrażliwości bialka na kardenolidy, zaobserwowano u 14 roznych gatunkow rownolegle substytucje w polipeptydach – to znaczy zamiany jednych rodzajow aminokwasów na inne w wyniku zamiany jednego nukleotydu na inny w DNA, wywolanej mutacja związane sa ze specjalizacja danego gatunku w odzywianiu sie roslinami produkujacymi kardenolidy.
Ponadto zaobserwowano cztery niezależne duplikacje genu kodujacego ATPaze (ATPa), w wyniku zbieznych wzorow ekspresji genow specyficznych dla jednego rodzaju tkanki.
Stwierdzamy zwiazek unikalnych dla tych grup owadow zbieznych podstawien aminokwasow z niedawnymi duplikacjami – co oznacza, ze zbieżna ewolucja dostosowujaca te owady do toksycznego pokarmu nie ciagnela sie przez niezliczone lata, tylko adaptacje nastapily niedawo, w szybszym tempie niz sie zaklada dla zmian w ramach ewolucji biologicznej (syntetycznej teorii ewolucji).”
Bywa raz tak, a raz inaczej, zmienność bez zmian w genomach:
https://academic.oup.com/mbe/article/37/6/1604/5728642
"Istnieje wiele przekonujących przykładów zbieżności molekularnej poszczególnych genów. Jednak częstość występowania i względne znaczenie adaptacyjnej konwergencji całego genomu pozostają w dużej mierze nieznane. Wiele ostatnich prac donosiło o uderzających przykładach nadmiernej konwergencji całego genomu, ale niektóre z tych badań zostały zakwestionowane z powodu użycia niewłaściwych modeli zerowych.
Tutaj zsekwencjonowaliśmy i porównaliśmy genomy 12 gatunków jaszczurek, które niezależnie zbiegły się w zestawach adaptacyjnych cech behawioralnych i morfologicznych. Pomimo szeroko zakrojonych poszukiwań sygnatury konwergencji molekularnej obejmującej cały genom, nie znaleźliśmy dowodów potwierdzających konwergencję molekularną określonych aminokwasów ani w poszczególnych genach, ani w porównaniach w całym genomie; nie odkryliśmy również żadnych dowodów potwierdzających nadmiar adaptacyjnej konwergencji w szybkościach podstawień aminokwasów w genach. Nasze odkrycia wskazują, że kompleksowa konwergencja fenotypowa nie znajduje odzwierciedlenia na poziomie kodowania białek u anoli w całym genomie, a zatem ta adaptacyjna konwergencja fenotypowa prawdopodobnie nie jest ograniczana przez ewolucję wielu specyficznych sekwencji. białkowych lub struktur."
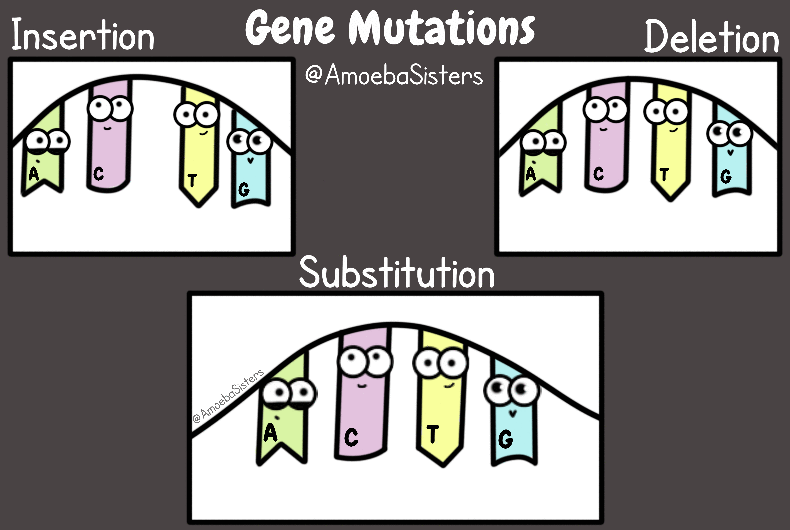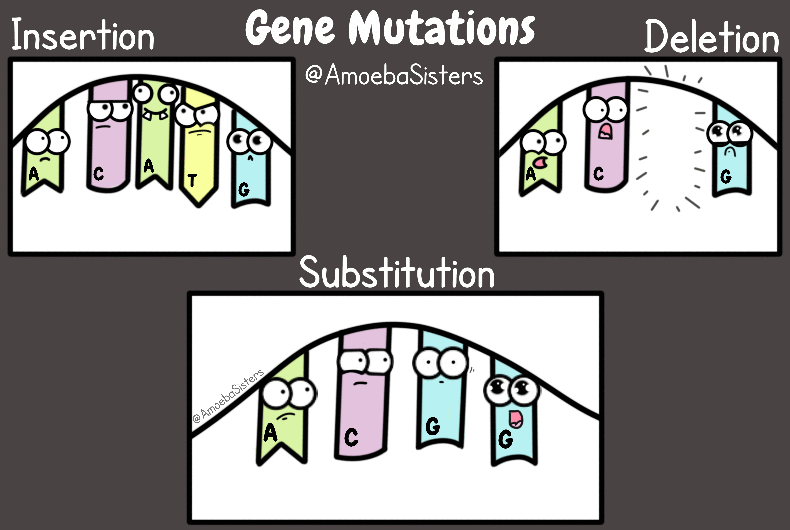
Darwiniści, jak i antydarwiniści popełniają te same błędy - formułują ostateczne wnioski na zbyt małej ilości faktów, lub budują rzekomo rzetelne tezy na hipotezach które domagają się uzasadnienia. Jak czytać literaturę naukową. Najlepiej zaczynać od prac przeglądowych :)
Np. przypadek prawa Dolla:
Prawo Dollo – hipoteza wysunięta przez francusko–belgijskiego paleontologa Louisa Dollo w roku 1890, mówiąca o nieodwracalności ewolucji.
Po raz pierwszy została sformułowana przez Dollo w brzmieniu „organizm nie jest w stanie powrócić nawet częściowo do formy uzyskanej przez przodków” (ang. An organism is unable to return, even partially, to a previous stage already realized in the ranks of its ancestors).
Według tej hipotezy struktura czy też organ utracony w procesie ewolucji nie pojawi się ponownie w linii organizmów, których przodkowie organ ów utracili. Wśród zwierząt znanych jest co najmniej kilka przykładów łamiących to prawo (choć nie wszyscy autorzy się z tym zgadzają), np. ponowne wykształcenie zębów żuchwowych u żab z rodzaju Gastrotheca przed 5–17 mln lat, po tym, jak płazy bezogonowe utraciły zęby żuchwy przed co najmniej 230 mln lat [za wikipedią]
http://bioblueprints.blogspot.com/2011/05/evolution-of-constraint-what-causes-and.html
„Kiedy organizmy tracą jakiś organ w ewolucji, rzadko można taką cechę ponownie odzyskać.
Pojęcie „Prawo Dollo” oraz definicja zostały po raz pierwszy sformułowane i rozpowszechnione w 1893 roku przez paleontologa Luisa Dollo.
Jednym ze znanych przykładów tej zasady jest liczba palców na tylnych i przednich kończynach kręgowców.
Niektóre z najwcześniejszych płazów kopalnych miały osiem, lub więcej palców u nóg, ale liczba ta została szybko zmniejszona do pięciu:
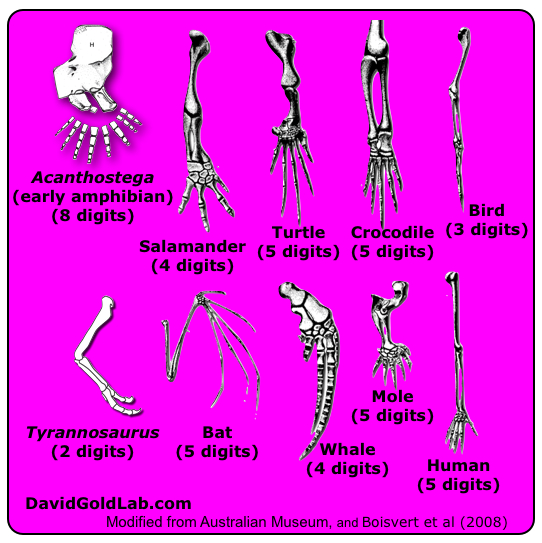
Przez ostatnie 300 milionów lat wszystkie kręgowce, od dinozaurów, ewolucyjnych przodków wielorybów po ludzi miały pięć lub mniej palców na przednich i tylnych kończynach, lub górnych i dolnych u H.sapiens. Ale dobrą zasadą jest to, że każde prawo filogenetyczne w biologii będzie miało wyjątki. Każdy, kto wie wystarczająco dużo o psach i kotach domowych wie, że niektóre rasy mają więcej palców. Oto łapa norweskiego Lundehunda. Posiada sześć palców:
Jaka przyczyna narusza prawo Dollo? Co w pierwszej kolejności wyjaśnia prawo Dollo? W następnych kilku postach zamierzam przyjrzeć się nauce stojącej za ograniczeniami ewolucyjnymi. Jakie przyczyny naruszają prawo Dollo?”

Porządny artykuł będący krytyczną oceną innego. Nie wiem na czym stanęło, bo nie śledziłem dalszych losów tej polemiki – każdy zainteresowany może sobie poszukać. Zwracam uwagę na końcowe wnioski streszczenia – autorzy zarzucają badaczom, których wnioski krytykują nieznajomość podstaw biologii ewolucyjnej, co skutkowało niechlujnym [ich zdaniem] materiałem rzekomo faktograficznym, jaki zaprezentowali. Kolejną lekcja: chcesz bronić, lub krytykować neodarwinowską wizję ewolucji i być poważnie traktowany, musisz zrozumieć postulowane przez darwinistów mechanizmy ewolucji.
Druga sprawa. Nie wystarczy przeczytanie jednego artykułu naukowego na jakiś ważny temat i na tej podstawie obalać przyjęte poglądy, tylko dlatego że został opublikowany w renomowanym czasopiśmie. Trzeba sprawdzić kontekst panujący na rynku idei naukowych, to znaczy: co na ten temat opublikowali inni badacze – wcześniej, lub ewentualnie później.
Kiedy w nauce pojawia się nowa koncepcja, to najpierw ma więcej przeciwników niż zwolenników i z czasem wywiązuje się ciekawa dyskusja. Nie na forach, tylko za pośrednictwem innych publikacji naukowych autorstwa fachowców z najwyższej półki! Po jakimś czasie nowa koncepcja zostaje zarzucona, potwierdzona lub pozostawiajaca dużo do zyczenia – najczęściej zostaje przy niej garść zwolenników, lub dzieli naukowców na dwa i więcej obozów.
Recenzenci przepuszczają dużo prac naukowych, ale to dopiero weryfikacja przez innych badaczy, na podstawie doświadczeń, stawia kropkę nad przysłowiowym „i”! Należy zwracać uwagę, czy badania są rzetelne, czy więcej jest sformułowań w rodzaju: „prawdopodobnie, przypuszczalnie, domaga się większej ilości eksperymentów”. Czy „wyniki potwierdzili inni badacze w wyniku ślepych prób, sprawa pewna bo wiele razy przetestowana [i tutaj odnośniki do odpowiednich prac]” i tak dalej. Kwestia prawa Dolla jest wieloznaczna i rozpatrywana w wielu aspektach w odniesieniu do opisów różnych zjawisk biologicznych i dlatego jeden tekst nie może obalać tej zasady, ale synteza wiedzy na ten temat pozwala wyrobić sobie obiektywną opinię.
Dlatego zapraszam do przeczytania tekstu o stworzeniach, które łamią prawo Dollo. Właściwie nie łamią tej samej zasady, którą na podstawie swojej skromnej wiedzy, w porównaniu z współczesną, formułował Dollo. On nie znał biologii molekularnej. Domaganie się od niego, gdyby żył, tłumaczenia się z jego wniosków, jest jak krytykowanie książki Karola Darwina „O pochodzeniu gatunków”, bo nic nie napisał o genach, czy biologii molekularnej w ogóle ????
https://onlinelibrary.wiley.com/doi/10.1111/j.1558-5646.2010.01041.x
„Z kilku ostatnich badań wynika, że wyjątki od uznanego za regułę prawa Dollo są częstsze niż dotychczas sądzono. Jeżeli te twierdzenia są prawdziwe, zmieniłoby to nasz pogląd na rolę ograniczeń rozwojowych planów ciała w ewolucji. Wyniki jednego z badań uczą, że nastąpiła reewolucja utraconych palców u jaszczurek z rodzaju Bachia. Oceniamy to twierdzenie.
Wnioskujemy, że zaproponowane molekularne drzewo filogenetyczne jest sprzeczne z mechanizmami ewolucyjnymi dotyczącymi biogeografii jaszczurek oraz z filogeniami opartymi na morfologii. Ponowna analiza danych molekularnych nie potwierdza topologii opublikowanego drzewa filogenetycznego tych gadów. Co więcej, dwa luźne założenia traktujemy jako nadinterpretacje, mianowicie że liczby palców są stałe i że polidaktylia ewoluuje niezależnie od innych parametrów morfologicznych.
Dochodzimy do wniosku, że nie ma przekonującego poparcia dla reewolucji liczby palców u Bachii. Omawiamy nasze ustalenia w świetle aktualnych dowodów na odwrócenie utraty cech złożonych. Dochodzimy do wniosku, że u metazoanów wyjątki od prawa Dollo znajdują się głównie wśród cech merystycznych, które powstają stosunkowo późno podczas embriogenezy. Kiedy systemy rozwojowe są bardziej skompartmentalizowane.
Wreszcie nasze badanie pokazuje, że analizy filogenetyczne powinny obejmować mechanizmy ewolucyjne, w tym ograniczenia, zmienność i selekcję. Nie tylko w celu prawidłowej rekonstrukcji filogenetycznej, ale także w celu prawidłowego wnioskowania ewolucyjnego.”
Inne spojrzenie:
O stworzeniach, które wystąpiły przeciwko ewolucyjnemu Prawu Dollo – wsteczna ewolucja, czy mechanizm w ramach normy reakcji na środowisko?
Żaby z rodzaju Gastrotheca po 200 000. 000 lat odzyskały zęby
https://onlinelibrary.wiley.com/doi/full/10.1111/j.1558-5646.2011.01221.x
REEWOLUCJA UTRACONYCH ZĘBÓW ŻUCHWYCH U ŻAB PO PONAD 200 MLN LAT I PONOWNA OCENA PRAWA DOLLO

„Prawo Dollo mówi, że struktury utracone ewolucyjnie nie już powrócą. Ostatnie analizy filogenetyczne ujawniły kilka potencjalnych przykładów, w przypadku których prawo Dollo wydaje się nie mieć zastosowania. Jednak te przykłady były ostatnio kwestionowane na podstawie zarzutów dotyczących błędów metodologicznych. W tym artykule dokumentuję uderzający i niepodważalny przykład reewolucji utraconej, złożonej struktury: zębó w żuchwie u żab z rodzaju Gastrotheca.
Używam skalibrowanej czasowo analizy filogenetycznej dla 170 gatunków płazów, aby wykazać, że zęby w żuchwie tych żab zostały utracone u wspólnego przodka współczesnych żab [nadinterpretacja, powinno być: wspólnego przodka obecnie żyjących żab z rodzaju Gastrotheca. Analiza ta nie wyjaśnia właściwości morfologicznych i strategii adaptacyjnych wszystkich współczesnych żab! Aż kusi żeby opisać te cuda, a właściwie przepisać ???? ]
Co najmniej 230 milionów lat temu i potem w historii geologicznej zęby były odzyskiwane. Ostatnio w ciągu ostatnich 5–17 milionów lat. Przeglądałem ostatnie badania nad reewolucją cech morfologicznych i na tej podstawie wykazuję, że ten długi okres braku cechy, jaką jest uzębiona szczęka i ponowne nabycie tej właściwości morfologicznej jest w dużej mierze bezprecedensowy.
Twierdzę również, że istnieje kilka problemów metodologicznych, które mogą powodować, że reewolucja cech będzie bardzo trudna do wykrycia w warunkach najbardziej prawdopodobnych, co prowadzi do błędnych interpretacji wiodących do podtrzymywania [neodarwinowskiej idei] prawa Dolla. Na koniec omówię możliwe mechanizmy, które mogą ułatwić ponowną [nielosową] ewolucję cech, które zanikły, w tym ewolucję zębów w żuchwie u żab, jako przykłady ograniczeń rozwojowych.”
https://www.nature.com/articles/nature01313

Przykład straszyków jest pierwszym, w którym udało się stwierdzić odzyskanie zdolności lotu po uprzedniej jego utracie. Do przypadków funkcji tak złożonych, jak zdolność lotu czy widzenie, stosuje się zwykle prawo nieodwracalności ewolucji. Mówi ono, że w procesie ewolucji niemożliwy jest powrót do wcześniejszego stadium rozwoju – nawet, gdy organizmy znajdują się w identycznych warunkach, jak ich przodkowie. Jeżeli np. któryś z narządów zaniknie, a na jego miejsce pojawi się podobny, to pochodzenie tych narządów jest inne. Zdaniem badacza, wyniki te mogą pociągnąć za sobą potrzebę zmian tego elementu teorii ewolucji. „To pierwszy przykład cechy złożonej, utraconej i odzyskanej później w trakcie ewolucji” – mówi Whiting. Informacje na temat wytworzenia skrzydeł (nawet, jeśli w danym momencie owad ich nie ma) zachowują się w genomie. Zdaniem Whitinga, genetyczne „instrukcje” wzrostu skrzydeł są podobne do „instrukcji”, według której wykształcają się odnóża. W długiej perspektywie czasu mogą być one „włączane” bądź „wyłączane” – podejrzewa badacz. Około 50 mln lat po utracie skrzydeł z jakiegoś powodu okazało się korzystne, by niektóre gatunki straszyków skrzydła odzyskały” – pisze Whiting. Obecnie znamy różne gatunki tych owadów – skrzydlate i bezskrzydłe.
Sprzeciw wśród darwinistów!
https://onlinelibrary.wiley.com/doi/full/10.1111/j.0307-6970.2004.00251.x
„W ostatnim numerze Nature , Whiting i inni (2003 ) donoszą o zaskakującym i nowatorskim wyniku badań dotyczących możliwości reewolucji owadzich skrzydeł od bezskrzydłych przodków u Phasmatodea. Jeśli to prawda, oznacza to ważny postęp w naszym zrozumieniu ewolucji owadów i genów. Odkrycie kwestionuje rzekomo od dawna utrzymywany pogląd, że skrzydła u owadów wyewoluowały tylko raz i były wielokrotnie bezpowrotnie tracone (na zasadach prawa Dolla). Jak twierdzą badacze ponownie wyewoluowane skrzydła mają dokładnie taką samą strukturę, jak inne skrzydła owadów. Gdyby ta cecha rzeczywiście została przywrócona u patyczaków, wykazałoby to niezwykłą stabilność genów kodujących rozwoj skrzydeł, pomimo inaktywacji tej ścieżki rozwojowej przez długie okresy czasu [jak u uzębionej żaby]. Wnioski Whitinga i innych zostały już opublikowane w literaturze popularnonaukowej ( New Scientist , Scientific American , The New York Times , Anon), ale zanim ten niezwykły ewolucyjny scenariusz trafi do podręczników entomologii należy je ponownie przeanalizować z zastosowaniem metody naukowej.
Dla hipotezy Whitinga i innych o reewolucji skrzydeł istotne są dwie linie dowodowe :
1. Rozklad na drzewie filogenetycznym patyczaków,
2. W tym rozkład transformacji skrzydel u osobnikow bez skrzydeł i uskrzydlonych na tym drzewie.
Ta ostatnia kwestia jest poruszona w niniejszym tekście. Nasza ponowna ocena materiału dowodowego przekonuje nas, że Whiting i inni znacznie przeszacowali prawdopodobieństwo reewolucji skrzydeł u patyczaków. Biorąc to pod uwagę nie widzimy podstaw do obalenia tradycyjnego poglądu na ewolucję patyczaków [i skrzydeł].
Czy jest możliwa reewolucja skrzydeł?
https://www.sciencedirect.com/science/article/pii/S0960982203003646
„Ewolucja skrzydeł jest powszechnie uważana za główny czynnik przyczyniający się do sukcesu owadów, dający wiele nowych możliwości rozprzestrzeniania się, radiacji adaptacyjnej, chwytania pożywienia i unikania drapieżników. Skrzydła owada (podobnie jak oczy) są przykładem złożonych struktur, które tak niepokoiły Karola Darwina i które pozostają ważnymi punktami badań nad którymi koncentrują się biolodzy rozwoju i biolodzy ewolucyjni.
Głównym pytaniem dotyczącym tych adaptacji strukturalnych – i innych złożonych cech, takich jak rozmnażanie płciowe – jest to, czy linie które je utracą, mogą je odzyskać. Przeważa pogląd, że taka reewolucja jest mało prawdopodobna, ponieważ po ich utracie geny wymagane do ich rozwoju powinny mieć możliwość akumulacji mutacji, a tym samym stać się niefunkcjonalnymi pseudogenami. Zagadnienie to zostało omówione bezpośrednio w niedawnej pracy Whitinga i innych: O ewolucji skrzydeł patyczaków (Phasmatodea). [….]”
Możliwości odzyskiwania oczów dowiedziono na przykładzie ryb jaskiniowych:
https://slawekp7.wordpress.com/2021/06/05/nielosowe-mutacje-w-goracych-miejscach-mutacyjnych-mechanizmy-epigenetyczne-ewolucja-w-super-tempie-o-nowej-syntezie-ewolucyjnej/
„[….] W przypadku dowiedzionego ponownego pojawienia się skrzydeł pouczające byłoby zbadanie podstaw ich rozwoju na poziomie molekularnym w różnych liniach uskrzydlonych gatunków – z uwzględnieniem stopnia pokrewieństw badanych grup owadów. Badanie powinno być przeprowadzone w celu ustalenia, czy wszystkie elementy mechanizmu rozwojowego przodków, u których zanikły skrzydła w wyniku dezaktywacji ścieżek rozwojowych, rzeczywiście zostały zachowane przez długie okresy czasu – co pozwoliło na ich uaktywnienie i ponowną morfogenezę skrzydeł.”
https://archive.nytimes.com/www.nytimes.com/learning/students/pop/20030122snapwednesday.html
„Naukowcom [neodarwinistom] trudno jest zaakceptować wyniki tych badań, ponieważ w ramach paradygmatu neodarwinowskiego utrzymuje się, że po utracie skrzydeł genetyczna maszyneria służąca do budowy tych organów nie powinna już podlegać doborowi naturalnemu, a co za tym idzie kumuluje mutacje niszczące cały ten molekularny system.
Niemniej jednak wynik rewolucyjnych badań przeszedł przez sito naukowych testów, a zespół rozszerzył swoje badania z 37 gatunków do 150! Autorzy zauważyli, że nowe badanie nie odpowiada na największe pytanie dotyczące tak złożonych cech, jak skrzydła. Chodzi o ich pochodzenie ewolucyjne w ogóle. Oznacza to, że żadne cykle utraty i odzyskiwania skrzydeł nie mogą wyjaśnić, w jaki sposób owady po raz pierwszy wyewoluowały swoje skrzydła [!!!].
Naukowcy stwierdzili, że odkrycia sugerują, że maszyneria genetyczna pozostała nienaruszona tych owadów prawdopodobnie dlatego, że bierze udział w morfogenezie innych narządów [efekt plejotropowy], jak nogi. Zespół zauważył, że to samo może dotyczyć innych maszynerii genetycznych odpowiedzialnymi za morfogenezę innych złożonych cech fenotypowych [np. utraconych i odzyskanych zębów u żab].
Jedyną grupą badaczy, która bez oporów przyjęła nowe odkrycie, są biolodzy rozwoju. Są przyzwyczajeni do radzenia sobie z mutantami, jak te z doświadczeń z muszkami owocowymi:noga wyrośnie zamiast czułka i vive versa.”
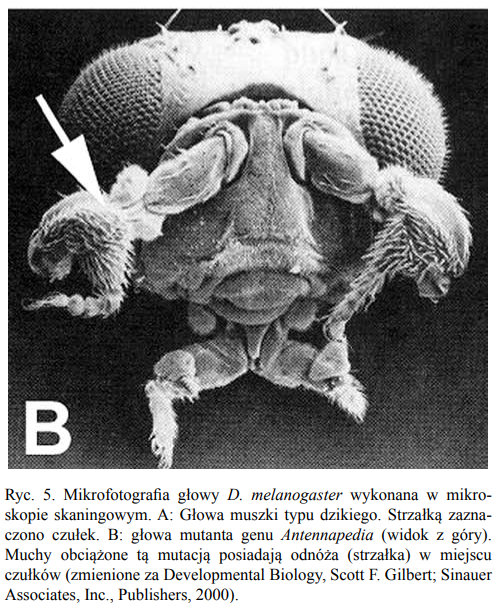
Jak nie argumentuje się na gruncie nauki - ewidentne przykłady!
Biologia teoretyczna (ewolucyjna) jest dobrym, uniwersalnym programem badawczym. Posługują się nim darwiniści i zwolennicy ID, ale wyciągamy różne wnioski. Od jakiegoś czasu, po mozolnym procesie obalania na podstawie dowodów tego, co zostało przyjęte bez dowodów pada jedna darwinowska ikona po drugiej. Tym razem nie dało się już dłużej reanimować koncepcji Samolubnego Genu Richarda Dawkinsa. Ma to ogromne konsekwencje dla darwinizmu, bo ta książka jest zlepkiem podstawowych koncepcji, na których opiera się tzw. syntetyczna teoria ewolucji (nowy neodarwinizm). Autorem tych hipotez nie jest Dawkins, tylko tytani biologii ewolucyjnej: John Maynard Smith, Hamilton, Edwards i inni. Antoni Hoffman w przedmowie do Ślepego Zegarmistrza Richarda Dawkinsa:
 Richard Dawkins ubrał wszystko w piórka własnego nazewnictwa i w stylu wprawnego gawędziarza przelał wszystko na papier – używając wprawnego pióra. Na pogrzebie hipotezy głównym mówcą był profesor Golik. Stwierdził, że zostają z nami podstawy: mutacje i dobór naturalny, który w jakiś sposób oddziałuje na geny nie gen. Że osobiście jest przekonany: za 20, 30, 40 lat będziemy wiedzieć więcej. Myślałem, że nauka bazuje na tym co wiemy a nie czego się być może dowiemy w przyszłości ?
Richard Dawkins ubrał wszystko w piórka własnego nazewnictwa i w stylu wprawnego gawędziarza przelał wszystko na papier – używając wprawnego pióra. Na pogrzebie hipotezy głównym mówcą był profesor Golik. Stwierdził, że zostają z nami podstawy: mutacje i dobór naturalny, który w jakiś sposób oddziałuje na geny nie gen. Że osobiście jest przekonany: za 20, 30, 40 lat będziemy wiedzieć więcej. Myślałem, że nauka bazuje na tym co wiemy a nie czego się być może dowiemy w przyszłości ?

„Samolubny gen” prof. Richarda Dawkinsa czy „Samolubne geny” prof. Pawla Golika?
https://slawekp7.wordpress.com/2021/02/12/gggggggo/
Zacytuję większy fragment wywodu Pawła Golika:
Z KONTEKSTEM
Biolog profesor Paweł Golik – DEBATA: Więcej niż samolubny gen – gen w dialogu ze środowiskiem” – debata XXII FN – 22.09.2018
„Geny tworzą cechy wpółpracując ze sobą – w żargonie nazywamy to oddziaływaniem epistatycznym. I trzeba to uwzględniać, dlatego że działanie doboru naturalnego na gen zależy nie tylko od wariantu genu, ale od całego jego otoczenia w genomie – mówimy wówczas o tle genetycznym. (…) Dlatego zamiast mówić o samolubnym genie należałoby mówić o samolubnych genach, samolubnych systemach, samolubnych genotypach, samolubnych genomach. Dobrze rozumiemy jak jest zbudowany gen i jak jest realizowany w komórce. Mało wiemy o wyższych poziomach interakcji genów. (…) Można to przyrównać do rozumienia czym jest cegła i poznawania czym cała budowla, która jest tym wyższym poziomem odnośnie którego teoria ewolucji będzie się musiała dużo dowiedzieć, jak dobór naturalny działa na geny. Jak ewolucja działa na pojedyńczy gen jest [hipotetycznie] dobrze opisane, jak na interakcje w sieciach genetycznych jest złożonym problemem, choć idea jest ta sama. (…)
[Epigenetyka] jest ciekawym rozbudowaniem tego, co wiemy o genetyce – tego jak geny są regulowane, ale to cały czas są geny, a właśnie zmiany (mutacje) w genach są podstawowym mechanizmem ewolucji. Natomiast to, jak zmiany w genach przekładają się na fenotyp organizmu jest procesem bardzo skomplikowanym i dopóki do końca nie zrozumiemy istyty tych mechanizmów nie będziemy do końca rozumieli jak działa ewolucja.
Dobór naturalny nie odczytuje genów, on działa na kodowane wielogenowo cechy fenotypowe i właśnie w tym aspekcie musimy się bardzo dużo dowiedzieć. Osobiście jestem przekonany, że teoretycy ewolucji za 10, 20, 30 lub 40 lat będą wiedzieć na ten temat znacznie więcej. [(neo)arwinizm] nadal będzie teorią, której rdzeń zasadniczo pozostanie ten sam – działanie doboru naturalnego przy udziale losowych czynników na losową zmienność genetyczną. Natomiast poznanie tego, jak współpracujące ze sobą geny wpływają na fenotyp i jakie są interakcje ewolucyjne fenotypu ze środowiskiem jest kwestią przyszłości – teoria ewolucji ma charakter rozwojowy. Racjonalnej alternatywy dla niej na horyzoncie jak nie było, tak nie ma!”
Niektóre definicje błędów logicznych popełnianych w dyskusjach


Logika odkrycia naukowego
https://pl.wikipedia.org/wiki/Logika_odkrycia_naukowego
"Logika odkrycia naukowego – książka wiedeńskiego profesora logiki i metodologii nauk i doktora psychologii, Karla Poppera, wydana w roku 1934, zawierająca nowatorskie jak na swoje czasy propozycje metodologiczne.
Jest to jedna z najważniejszych prac dotyczących filozofii nauki.
Popper uznaje dedukcję za racjonalny odpowiednik indukcji. Biorąc pod uwagę naukowy punkt widzenia, geneza hipotez generowanych na podstawie dedukcji jest nieistotna (antypsychologizm), to powody, dla których uznajemy je za naukowe są ważne. Tutaj Popper proponuje wykorzystać metodę falsyfikacji:
Propozycja ogólnej hipotezy.
Wydedukowanie z niej szczegółowych zdań empirycznych, tzw. zdań bazowych.
Doświadczalne sprawdzenie, czy zdania pokrywają się z rzeczywistością.
Hipotezę należy odrzucić, gdy zdania z niej wynikające okażą się fałszywe. W innym przypadku można tymczasowo uznać hipotezę za sprawdzoną oraz przyjąć jako naukową, co jednak nie zwalnia z przeprowadzania dalszych, surowszych jej prób.
Popper jako najcenniejsze widzi hipotezy ryzykowne, które poprzez precyzję obejmują szeroki zakres rzeczywistości. Nie nadaje im to jednak cech pewności – ostatecznym celem nauki jest bowiem eliminacja fałszu.
Popperowski model opisuje logikę, charakteryzującą naukę w sposób synchroniczny, wyznaczającą determinanty w jej rozwoju. Jest to model racjonalistyczny i autonomiczny, niezależny od zewnętrznych czynników socjologicznych, historycznych oraz psychologicznych."

Bez stawiania założeń i ich testowania nie można przeprowadzić procesu dowodowego, ale założenia (hipotezy), to nie dowody, a nawet nie teorie. Teorię naukową tworzy przynajmniej kilka dobrze potwierdzonych, pasujących do siebie hipotez opisujących genezę jakiegoś zjawiska biologicznego. W zależności od ilości i jakości potwierdzonych założeń mamy słabą, lub dobrą teorię wyjaśniająca. Żeby teorię można było nazywać faktem zbiór zgromadzonych i pozytywnie zweryfikowanych hipotez musi kreślić ostry obraz wyjaśniający.
Proces przypomina szybę pokrytą zmywalną farbą zasłaniającą duży namalowany obraz. Każda zweryfikowana hipoteza to jedno mniejsze, lub większe kółko powstałe w wyniku zmycia farby w określonym obszarze obrazu. Początkowo pozwala nam to widzieć fragmenty dzieła i stawiać słabe teorie lub alternatywne do nich. Nie można jeszcze poskładać wszystkich potrzebnych elementów do kupy, więc ostatecznego, albo najbardziej prawdopodobnego wyjaśnienia nie ma. Stanie się to dopiero wtedy, gdy miejsc ze zmytą farbą będzie tyle, żeby móc przeprowadzić spójną logicznie syntezę zdobytej wiedzy stwierdzając, co jest namalowane na obrazie.
Okazuje się, że namalowana jest maszynka do mielenia mięsa i to twierdzenie jest prawdziwe, ale pozostaje do wyjaśnienia, czy mielenie jest napędzane ręczną korbą czy silnikiem elektrycznym. Żeby to sprawdzić uczeni dalej projektują i prowadzą badania na podstawie stawianych i weryfikowanych hipotez – starając się zmyć farbę z reszty obrazu. Niemniej mamy już do czynienia z teorią i zarazem prawdą, faktem naukowym. Teraz chodzi jedynie o dopracowanie szczegółów, ale nawet jakby się nie udało, to nie zmieni to faktu, że na obrazie jest namalowana maszynka do mielenia mięsa. Zatem teoria naukowa nie musi być kompletna, żeby można było ją traktować jako prawdziwe, rzetelne wyjaśnienie na gruncie nauki. Nikt nie wątpi, że istnieje grawitacja.



Koncepcja naukowa, jako fakt i teoria
Newton sprawdził to doświadczalnie – podrzucił jabłko do góry a siła ciążenia spowodowała, że spadło na ziemię. Niemniej fizycy po dziś dzień nie mają pojęcia, jakie zjawiska kwantowe są odpowiedzialne za istnienie grawitacji. Zatem w obrębie badań nad grawitacją napotykamy twarde fakty i hipotezy – jeżeli w ogóle istnieją warte poważnego rozważenia. Być może nigdy nie dowiemy się, jakie czynniki są odpowiedzialne za działanie grawitacji, ale czy ktoś rozsadny z tego powodu zaneguje jej istnienie? Neuronaukowcy ponad wszelką wątpliwość udowodnili, że mózg jest odpowiedzialny za świadomość, czy to w wyniku myślenia czy przetwarzania sygnałów napływajacych z otoczenia za pośrednictwem narzadów zmysłów. Istnieje jakiś neurobiolog, który potrafi opisać mechanizmy w komórkach nerwowych, których współpraca pozwala ludziom i zwierzętom odczuwać świadome doznania? Komórki nerwowe, neurony czy glejowe, tworzace mózg składają się, jak wszystkie inne rodzaje komórek z łańcuchów biologicznych: DNA, RNA, z cukrów, polipeptydów, i z błon tłuszczowych.

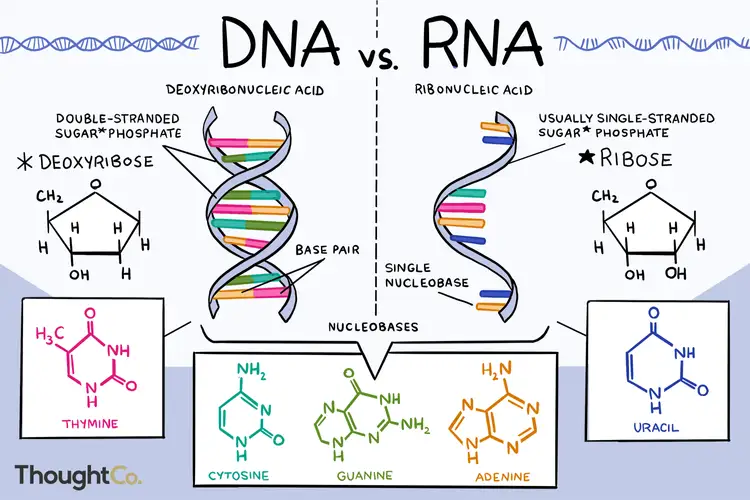
DNA, RNA, cukry i polipeptydy, które zwijają się w różne białka oraz błony komórkowe złożone są z mniejszych elementów. „DNA i RNA są polimerami (w przypadku DNA, często bardzo długimi polimerami), i są zbudowane z monomerów znanych jako nukleotydy. Kiedy połączymy te monomery, powstający łańcuch jest nazywany polinukleotydem (poly- = „wiele”). Każdy nukleotyd jest zbudowany z trzech części: pierścienia zawierającego azot, pięciowęglowego cukru i przynajmniej jednej grupy fosforanowej.” [1] Innymi słowy nukleotydy składają się z atomów.
https://slideplayer.pl/slide/10515460/
Polipepotydy z odpowiednio połaczonych aminokwasów. Aminokwasy składają się z atomów wodoru, azotu, wegla i tlenu [2]
Cukry (węglowodany, sacharydy) w ścisłym znaczeniu są to organiczne związki chemiczne składające się z atomów węgla oraz wodoru i tlenu.[3]
https://eszkola.pl/chemia/weglowodany-8475.html


Tłuszcze, lipidy tworzĄce błony komórkowe – składają się z atomów wodoru, węgla i tlenu [4] Podsumowując tą wiedzę rozumiemy, że mózg składa się z atomów, które tworzą monomery i polimery, a ich odpowiednia struktura i współpraca pozwala na świadome doznania.
Tłuszcze, lipidy tworzĄce błony komórkowe – składają się z atomów wodoru, węgla i tlenu [4] Podsumowując tą wiedzę rozumiemy, że mózg składa się z atomów, które tworzą monomery i polimery, a ich odpowiednia struktura i współpraca pozwala na świadome doznania.
https://portal.scholaris.pl/zasob/58708

Jak to się dzieje, ze ten system uporządkowanych atomów wywołuje subiektywne wrażenie Świadomości nie wie nikt. Ktoś na podstawie tej niewiedzy zaneguje fakt, ze to mózg jest odpowiedzialny za wiadomość? Czy ta luka w wiedzy przeszkadza w dalszym badaniu i poznawaniu funkcji mózgu? Gdyby powiększyć atom i umieścić w dużym pokoju, to gdyby jądro było po środku, to orbity elektronów znajdowałyby się w pobliżu ścian. Atom to w większości puste przestrzenie, złożony z atomów mózg również [5] Istnieją hipotezy, ze za Świadomość odpowiadają zjawiska kwantowe. Jeżeli to prawda, to nikt ich nie zbadał i nie potrafi opisać. [6]

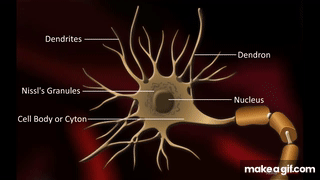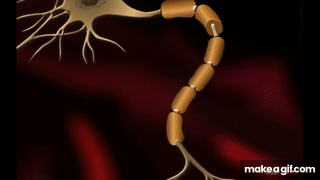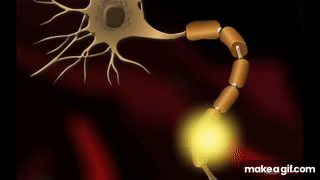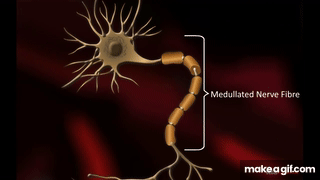

Oszałamiająca złożoność wynikająca z prostoty – wszystko, co żyje składa się z mikroskopijnych łańcuchów!
Biologia teoretyczna (ewolucyjna). Narzędzia badawcze i metody dochodzenia do prawdy – logika odkryć naukowych [7]




Proces przypomina powolne wznoszenie się podnośnika z obserwatorem. Najpierw widzi niewiele, ale niejako z lotu ptaka coraz więcej, aż w końcu granice jakiejś osady. Dopiero teraz może wykonywać liczne zdjęcia terenu i na ich podstawie kreslić wiarygodne mapy. Dochodzenie do poznania obiektywnej prawdy naukowej można porównać do pracy detektywa, który szuka i co jakiś czas znajduje poszczególne puzzle, a następnie stara się je połączyć ze znalezionymi wczesniej. Początkowo różne puzle dają fragmentaryczny obraz, ale z czasem niektore z nich zaczynają do siebie pasować i w tych miejscach układanka staje się coraz lepiej rozumiana. Wróćmy do metod badawczych, jakie stosują biolodzy teoretyczni.


W biologii pojęcie EWOLUCJA ma różne znaczenia:

https://arctic.au.dk/news-and-events/news/show/artikel/from-brown-to-white-evolution-of-the-polar-bear/
„Niedźwiedź polarny wyewoluował od niedźwiedzia brunatnego, zmieniając kolor futra na biały – idealny do wtapiania się w pokryte śniegiem i lodem środowisko. Dzięki nowym danym międzynarodowy zespół badawczy dowodzi, że oddzielenie niedźwiedzia polarnego od brunatnego przebiegało bardzo szybko.
„Porównaliśmy geny niedźwiedzi brunatnych i polarnych i byliśmy zaskoczeni. Przede wszystkim niedźwiedź polarny jako podgatunek ma mniej niż 480 000 lat. Jest więc znacznie młodsza niż wcześniej sądzono” – mówi profesor Rune Dietz z Centrum Badań Arktycznych, Wydział Nauk Biologicznych Uniwersytetu w Aarhus.
Naukowcy są przekonani, że niedźwiedź brunatny migrował na północ podczas cieplejszego okresu klimatycznego, a gdy później nastał okres chłodu, grupa niedźwiedzi brunatnych prawdopodobnie została odizolowana i dlatego zwierzęta zostały zmuszone do szybkiego przystosowania się do nowych warunków.
Naukowcy pokazują również, że niedźwiedź polarny rozwinął zdolności do życia w Arktyce w ciągu zaledwie kilkuset tysięcy lat – a to bardzo szybka ewolucja.
(Impulsy ze środowiska), zmiany w genach (w ilościach kopii danego genu i wzorach ekspresji genów aktywowanych w odpowiedzi na nowe wyzwania otoczenia) i (być może) działania (w jakimś zakresie) doboru naturalny nie tylko doprowadziły do zmiany ubawienia sierści niedźwiedzia polarnego na biały kolor, ale dostosowały do nowej diety. Za te adaptacje odpowiadają zmiany w genach regulujących transport tłuszczu we krwi i rozkład tkanki tłuszczowej w organizmie.
Kiedy naukowcy porównują zestaw genów niedźwiedzi polarnych z ludzkimi, wyraźnie widoczne są zmiany w genie apoB. Jest to gen, który u ssaków koduje najważniejsze białko w LDL (lipoproteina o niskiej gęstości), powszechnie znane jako „zły” cholesterol.
Jest to bardzo ważna adaptacja do życia w rejonach Arktyki, gdzie niedźwiedzie polarne głównie żywią się mięsem bogatym w tłuszcz pozyskiwanym z upolowanych ssaków morskich, przystosowanych do życia w ekstremalnie niskich temperaturach.
Dieta niedźwiedzi polarnych jest zdominowana przez tłuszcze, jednak w ich tętnicach nie ma złogów (blaszki miażdżycowej), dzięki czemu unikają one chorób sercowo-naczyniowych.
„Zmiany w tym genie mogą decydować o tym, że niedźwiedź polarny radzi sobie z wysokimi stężeniami tłuszczu, cholesterolu i cukru we krwi” – wyjaśnia profesor i lekarz weterynarii Christian Sonne z Centrum Badań Arktycznych, Wydziału Bioscience, Uniwersytet w Aarhus.”:
Link kieruje do oryginalnej pracy:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6613075/

W ramach uzupełnienia argumentacji zawartej w powyższym streszczeniu, oryginalnie pokaleczonym neodarwinowskimi określeniami, które nie maja nic wspólnego z opisywanym zjawiskiem, ale za to potrafią namieszać w głowach ludziom, którzy nie maja odpowiedniej wiedzy (w nawiasach zawarłem sprostowania).
Skomentowałem jeden z wywodów właściciela kanału „Naukowy bełkot”, który nie mając pojęcia o czym mówi zamienił swój wywód w pseudo-naukowy bełkot! ?
Mieszając pojęcia, procesy i postulowane mechanizmy adaptacje w ramach normy reakcji na środowisko zaprezentował, jako pięć dowodów na losową ewolucję w ujęciu tzw. syntetycznej teorii ewolucji (współczesnego neodarwinizmu). Np. Zięby Darwina z wysp Galapagos. Po wypowiedzi właściciela kanału „Naukowy bełkot” wynika, że nie jest on zapoznany z wynikami najnowszych badań nad genetyką i zmiennością tych ptaków. Sam dał się oszukać i sprzedaje to dalej i jeszcze mu za to płacą:
Zięby Darwina, to jedna z ikon darwinizmu, jednak mało kto spoza grona fachowców i osób szerzej zainteresowanych nauką wie, że co do tego wniosku nie ma zgody wśród naukowców. Ptaki te są uważane za podręcznikowy przykład radiacji adaptacyjnej, w wyniku … Czytaj dalej →
https://slawekp7.wordpress.com/2018/11/13/zieby-darwina-jeden-gatunek-czy-wiele/
Źródła:
[A]
ps://pl.wikipedia.org/wiki/Dob%C3%B3r_naturalny
https://zpe.gov.pl/a/zmiennosc-organizmow/D1B3tf0YZ
[B]
Hipotezy i teorie naukowe o różnym stopniu wiarygodności
https://pl.wikipedia.org/wiki/Hipoteza
https://pl.wikipedia.org/wiki/Teoria_naukowa
https://pl.wikipedia.org/wiki/Rodzaje_twierdze%C5%84_naukowych
Materiały źródłowe dotyczące wstępu:
[1]
https://pl.khanacademy.org/science/ap-biology/gene-expression-and-regulation/dna-and-rna-structure/a/nucleic-acids
[2]
https://pl.wikipedia.org/wiki/Aminokwasy
[3]
https://pl.wikipedia.org/wiki/W%C4%99glowodany
[4]
https://pl.wikipedia.org/wiki/T%C5%82uszcze
[5]
Polecane
Regulacja ekspresji genu
https://slideplayer.pl/slide/2868214/
https://www.ucsf.edu/news/2017/02/405686/mysterious-98-scientists-look-shine-light-our-dark-genome
Tajemnicze 98% ludzkiego genomu: naukowcy chcą rzucić światło na tą część ludzkiego genomu, która jest przyrównywana do ciemnej materii
Po zakończeniu w 2003 roku Projektu Ludzkiego Genomu – który zsekwencjonował wszystkie 3 miliardy „ liter ” lub par zasad w ludzkim genomie – wielu myślało, że nasze DNA stanie się otwartą księgą. Szybko jednak pojawił się kłopotliwy problem: chociaż naukowcy mogli dokonać transkrypcji książki, mogli zinterpretować tylko niewielki jej procent.
Tajemnicza większość – aż 98 procent – naszego DNA nie koduje białek. Uważa się, że większość tego „genomu ciemnej materii” to niefunkcjonalne resztki ewolucyjne, które dopiero czekają na przejażdżkę. Jednak wśród tego niekodującego DNA kryje się wiele kluczowych elementów regulacyjnych, które kontrolują aktywność tysięcy genów. Co więcej, pierwiastki te odgrywają ważną rolę w chorobach, takich jak rak, choroby serca i autyzm, i mogą stanowić klucz do możliwych wyleczeń.
Dowód na jednorazową utratę zmineralizowanych zębów u wspólnego przodka ptaków
https://science.sciencemag.org/content/346/6215/1254390
Struktura i funkcja ameloblastyny jako białka macierzy zewnątrzkomórkowej: adhezja, wiązanie wapnia i interakcja z CD63 u ludzi i myszy
Ludzie i ptaki mają te same geny odpowiedzialne za uczenie się mowy i śpiewu
https://www.newscientist.com/article/dn26694-humans-and-birds-share-the-same-singing-genes/#ixzz72deMHAsk
Analiza genomowa ujawnia plejotropowe allele w EDN3 i BMP7 zaangażowane w kolor grzebienia kurczaka i produkcję jaj
Identyczne krótkie sekwencje peptydowe w niepowiązanych białkach mogą mieć różne konformacje: poligon testowy dla teorii rozpoznawania immunologicznego
Ewolucja zbieżna w elementach strukturalnych białek badanych za pomocą analizy profili krzyżowych
https://pubmed.ncbi.nlm.nih.gov/22244085/
Białka o tej samej fałdzie i niepowiązanych sekwencjach mają podobny skład aminokwasowy
https://pubmed.ncbi.nlm.nih.gov/16565950/
Głęboka homologia w erze sekwencjonowania nowej generacji
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5182409/
Jak różne sekwencje aminokwasów determinują podobne struktury białek: Struktura i dynamika ewolucyjna globin
https://www.sciencedirect.com/science/article/abs/pii/0022283680903733
Wykrywanie adaptacyjnej, zbieżnej ewolucji aminokwasów
https://royalsocietypublishing.org/doi/10.1098/rstb.2018.0234
W genomice ewolucyjnej naukowcy zainteresowali się identyfikacją substytucji, które przyczyniają się do zbieżnych adaptacji fenotypowych. Jest to trudne pytanie, które wymaga odróżnienia zbieżnych podstawień pierwszego planu, które są zaangażowane w zbieżny fenotyp od tła zbieżnego podstawienia. Mogą być one powiązane z innymi adaptacjami, mogą być neutralne lub mogą być konsekwencją błędów mutacyjnych. Ponadto nie ma ogólnie przyjętej definicji zbieżnych podstawień. W literaturze zaproponowano różne metody, które wykorzystują różne definicje, co skutkuje różnymi zestawami kandydujących zbieżnych podstawień pierwszego planu. W tym artykule najpierw opisujemy procesy, które mogą generować zbieżne podstawienia pierwszego planu w sekwencjach kodujących, oddzielając procesy adaptacyjne od nieadaptacyjnych. Po drugie, dokonujemy przeglądu metod, które zostały zaproponowane do wykrywania zbieżnych podstawień pierwszego planu w sekwencjach kodujących i ujawniają założenia, które leżą u ich podstaw.
FOXP2
https://www.pbs.org/wgbh/nova/teachers/activities/0304_01_nsn.html
Koncepcja ewolucyjna w genetyce i genomice
https://www.ncbi.nlm.nih.gov/books/NBK20255/
https://academic.oup.com/mbe/article/37/6/1604/5728642
Istnieje wiele przekonujących przykładów zbieżności molekularnej poszczególnych genów. Jednak częstość występowania i względne znaczenie adaptacyjnej konwergencji całego genomu pozostają w dużej mierze nieznane. Wiele ostatnich prac donosiło o uderzających przykładach nadmiernej konwergencji całego genomu, ale niektóre z tych badań zostały zakwestionowane z powodu użycia niewłaściwych modeli zerowych. Tutaj zsekwencjonowaliśmy i porównaliśmy genomy 12 gatunków jaszczurek, które niezależnie zbiegły się w zestawach adaptacyjnych cech behawioralnych i morfologicznych. Pomimo szeroko zakrojonych poszukiwań sygnatury konwergencji molekularnej obejmującej cały genom, nie znaleźliśmy dowodów potwierdzających konwergencję molekularną określonych aminokwasów ani w poszczególnych genach, ani w porównaniach w całym genomie; nie odkryliśmy również żadnych dowodów potwierdzających nadmiar adaptacyjnej konwergencji w szybkościach podstawień aminokwasów w genach. Nasze odkrycia wskazują, że kompleksowa konwergencja fenotypowa nie znajduje odzwierciedlenia na poziomie kodowania białek u anoli w całym genomie, a zatem ta adaptacyjna konwergencja fenotypowa prawdopodobnie nie jest ograniczana przez ewolucję wielu specyficznych sekwencji białkowych lub struktur.
https://www.sciencedirect.com/topics/medicine-and-dentistry/convergent-evolution
”Ewolucja zbieżna odnosi się do ewolucji w różnych rodach struktur, które są podobne lub „analogiczne”, ale nie można tego przypisać istnieniu wspólnego przodka; innymi słowy, fakt, że struktury są analogiczne, nie odzwierciedla homologii. Podobieństwo może leżeć na poziomie fenotypowym, w którym to przypadku linie mają tę samą jawną cechę, ale podstawowe sekwencje DNA są różne.
Ewolucja zbieżna zachodzi, gdy gatunki zajmują podobne nisze ekologiczne i adaptują się w podobny sposób w odpowiedzi na podobne presje selekcyjne. Cechy, które powstają w wyniku zbieżnej ewolucjisą określane jako „struktury analogiczne”. Kontrastowane są ze „strukturami homologicznymi”, które mają wspólne pochodzenie. Przeciwieństwem ewolucji konwergentnej jest ewolucja rozbieżna, w której spokrewnione gatunki rozwijają różne cechy.
Dobrze udokumentowane przypadki zbieżnej ewolucji podobnych sekwencji DNA nie są liczne; takie przypadki są zwykle ograniczone do kilku aminokwasów. Ewolucja zbieżna może wprowadzać w błąd biologów zajmujących się filogenezą, ponieważ naśladuje wspólne pochodzenie. Standardowe metody filogenetyczne nie są przystosowane do rozróżnienia między nimi. Kiedy ewolucja zbieżna jest mylona z homologią, powstaje drzewo filogenetyczne, które ma zafałszowany wygląd, to znaczy gatunki wydają się pochodzić od wspólnego przodka, podczas gdy w rzeczywistości tak nie jest.”
https://akronfossils.wordpress.com/2012/11/13/truth-of-junk-dna-verified/
https://www.nature.com/articles/35057062
„Chociaż wyniki ENCODE sugerują, że co najmniej 80% „śmieciowego” DNA faktycznie odgrywa rolę w kontrolowaniu innych genów, Birney uważa, że bardziej prawdopodobne jest, że prawie 100% z niego okaże się mieć cel, a wiele innych genów regulacyjnych czeka na odkrycie. ”
https://geneticliteracyproject.org/2018/01/08/junk-dna-mining-genomes-dark-matter-new-disease-treatments/
Pilnie potrzebne są zmiany paradygmatu w poszukiwaniu nowych leków, ponieważ tradycyjna ścieżka odkrywania leków prawie się wyczerpała. Być może w przyszłości sektor farmaceutyczny wykorzysta unikalne mocne strony kilku strategii do opracowania rozwiązań kombinatorycznych. Tylko czas pokaże, czy rozwiązania terapeutyczne oparte na „śmieciowym DNA” przetrwają próby kliniczne. Pomysł wydaje się jednak nowatorski i bardzo interesujący, by zasługiwał na bliższe przyjrzenie się.
Zakres funkcjonalności w ludzkim genomie
John S Matticki
Marcel E Dinger
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4685169/
https://europepmc.org/article/med/25699073
https://wp-projektu.pl/nowosci/nature-reviews-genetics-funkcjonalnosc-pseudogenow-przedwczesnie-odrzucona/
Nowy artykuł w „Nature Reviews Genetics”, Overcoming Challenges and Dogmas to Understand the Functions of Pseudogenes1 [Pokonywanie wyzwań i dogmatów w celu zrozumienia funkcji pseudogenów], jest po prostu niesamowity. Dokumentuje nie tylko, że pseudogeny pełnią ważne funkcje, ale także że ze względu na obecne „dogmaty” w biologii, jak również na współczesne ograniczenia techniczne ‒ nie rozpoznajemy ich prawdziwych funkcji. Jak ujęli to Seth Cheetham i współautorzy, biologia ucierpiała w wyniku „braku motywacji do badania funkcji pseudogenów z powodu przyjęcia założenia, że nie pełnią one żadnych funkcji”2, przy czym „dominujące ograniczenie w postępach badań pseudogenów ma związek z przekonaniem, że regiony pseudogenów są z natury niefunkcjonalne”3.
W streszczeniu artykułu autorzy bardzo jasno przedstawili swoją tezę:
Pseudogeny są zdefiniowane jako regiony genomu zawierające wadliwe kopie genów. Znajdujemy je w prawie wszystkich formach życia, a w genomach ssaków występują w podobnej liczbie, co rozpoznane geny kodujące białka. Chociaż często przypuszcza się, że nie pełnią one żadnych funkcji, okazuje się, że coraz więcej pseudogenów odgrywa ważne role biologiczne. Biorąc pod uwagę ich ewolucyjne pochodzenie i ograniczenia metod sekwencjonowania genomu, twierdzimy, że pseudogeny zostały błędnie sklasyfikowane. Naszym zdaniem powszechne nieporozumienie dotyczące pseudogenów, utrwalone częściowo przez przypisanie im pejoratywnego określenia „pseudogen”, doprowadziło do częstego wykluczania ich z badań funkcjonalności i z analiz genomowych. W naszej opinii wraz z pojawieniem się technik upraszczających badanie pseudogenów obiektywne spojrzenie na te elementy genomowe umożliwi ujawnienie cennych informacji na temat funkcji i ewolucji genomu4.
Zwracają oni uwagę, że wiele obszarów DNA, które pierwotnie uznano za pseudogenowe śmieci, odgrywa jednak określoną rolę: „zważywszy na to, że rośnie liczba regionów pierwotnie zidentyfikowanych jako pseudogenowe, które później okazały się pełnić funkcje biologiczne, istnieje ryzyko, że pewne regiony genomu są przedwcześnie odrzucane jako pseudogeny i uważane za pozbawione funkcji”5.
Szukajmy funkcji, a je znajdziemy [….]
Rozbieżność ewolucyjna i Konwergencja w białkach
1.

2.

3.

http://docplayer.pl/112542763-Budowa-biogeneza-i-mechanizm-dzialania-kompleksu-mitochondrialnej-syntazy-atp.html
"Biogeneza syntazy ATP jest procesem złożonym, wymaga bowiem udziału licznych białek niebędących podjednostkami enzymu, regulujących ekspresję genów syntazy oraz składanie podjednostek w dojrzały enzym. Niniejsze opracowanie stanowi podsumowanie aktualnego stanu wiedzy o budowie, biogenezie i mechanizmie działania kompleksu syntazy ATP."

4.









https://wp-projektu.pl/wp-content/uploads/2021/06/Behe_Precyzyjny.projekt.pdf










http://kosmos.icm.edu.pl/PDF/2018/195.pdf


https://www.kul.pl/files/57/pdf/sagan_spor.o.nieredukowalna.zlozonosc.pdf






----

 ------
------

























 ---
---































Komentarze