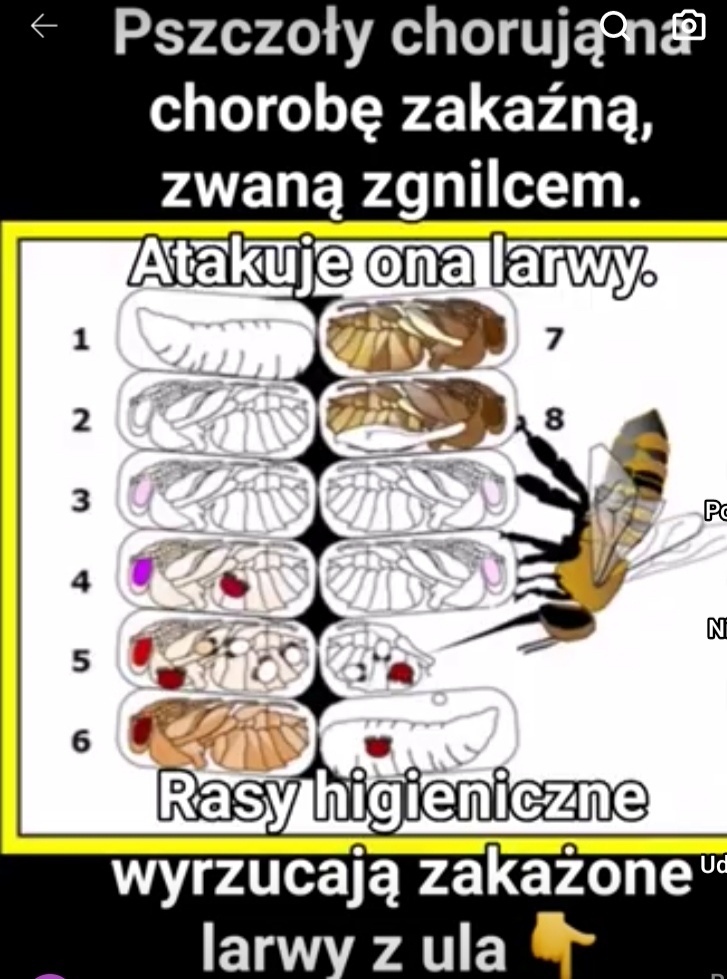
Pszczoły chorują na chorobę zakaźną, zwaną zgnilcem. Atakuje ona larwy w komórkach plastra. Wśród udomowionych ras pszczelich ryzyko zapadnięcia na tę chorobę jest u niektórych ras większe niż u innych i jak się okazuje różnica między szczepami – przynajmniej w niektórych przypadkach – przejawia się na poziomie behawioralnym. Istnieją szczepy zwane higienicznymi, które szybko opanowują epidemię, lokalizując zakażone larwy, a następnie wydobywając je z komórek i wyrzucając z ula.
„Pszczoły chorują na chorobę zakaźną, zwaną zgnilcem. Atakuje ona larwy w komórkach plastra. Wśród udomowionych ras pszczelich ryzyko zapadnięcia na tę chorobę jest u niektórych ras większe niż u innych i jak się okazuje różnica między szczepami – przynajmniej w niektórych przypadkach – przejawia się na poziomie behawioralnym. Istnieją szczepy zwane higienicznymi, które szybko opanowują epidemię, lokalizując zakażone larwy, a następnie wydobywając je z komórek i wyrzucając z ula.
Inne szczepy nie praktykują tej dzieciobójczej higieny i te podatne są na chorobę. Zachowanie zaangażowane bezpośrednio w zabiegi higieniczne jest dość złożone.Robotnice muszą zlokalizować komórkę z zakażoną larwą, usunąć z komórki woskową pokrywę, wyciągnąć larwę, przeciągnąć ją przez wejście do ula i zrzucić na stertę śmieci.

https://youtube.com/shorts/ABtMIkWEgp4?si=DaIFPpmVD3kzn7pv
Prowadzenie eksperymentów genetycznych na pszczołach jest z wielu powodów sprawą skomplikowaną. Robotnice z reguły się nie rozmnażają, krzyżuje się więc królową jednego szczepu z trutniem (czyli samcem) wywodzącym się z innego, a następnie obserwuje się zachowanie potomnych robotnic.
Tym właśnie zajmował się W. C. Rothenbuhler. Stwierdził on, że w pierwszym pokoleniu mieszańców wszystkie roje były niehigieniczne. Geny na higieniczność wciąż były obecne, ale okazały się recesywne. Gdy Rothenbuhler skrzyżował wstecznie mieszańce pierwszego pokolenia z czystym szczepem higienicznym otrzymał przepiękny rezultat.
Wśród potomnych rojów dało się wyróżnić trzy grupy. Jedna z grup demonstrowała pełny zestaw zachowań higienicznych, druga nie wykazywała ich w ogóle, trzecia natomiast zatrzymywała się w pół drogi. W grupie tej robotnice otwierały woskowe komórki z chorymi larwami, ale nie wyrzucały larw z ula.
Pszczoły higieniczne pozbawione funkcji jednego z genów kodujących zachowania higieniczne nie potrafią otwierać wieczka komory plastra, aby wydobyć z niej zakażoną larwę i wyrzucić ją z ula.
Rothenbuhler podejrzewał, że mogą istnieć osobne geny: na otwieranie komórek i na wyrzucanie larw.
U w pełni higienicznego szczepu jeden i drugi gen ulegały ekspresji spełniając swoje funkcje.
Mieszańce, które zatrzymywały się w pół drogi, przypuszczalnie posiadały aktywny gen na otwieranie komórek, ale gen na pozbywanie się zarażonych larw był wyciszony.
Rothenbuhler domyślał się, że grupa pszczół, która nie przejawiała zachowań higienicznych może posiadać aktywny gen na wyrzucanie, którego efekt nie mógł się przejawić z powodu braku genu na otwieranie. Potwierdził to bardzo eleganckim eksperymentem, w którym usunął pokrywy komórek osobiście.
Połowa spośród wcześniej niehigienicznych pszczół zaczęła przejawiać zahowania higieniczne. Gdy eksperymentator usunął przeszkodę odsłaniając zainfekowaną larwę owady podchodziły do otwartej komory, chwytały chorą larwę w żuwaczki i wyrzucały z ula. Gen na wyrzucanie jest bezużyteczny, jeśli nie towarzyszy mu aktywny gen na otwieranie i odwrotnie.”

Richard Dawkins, „Samolubny gen” strona 95
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4491870/
„Zachowanie higieniczne to złożona, uwarunkowana genetycznie cecha, która służy jako kluczowy mechanizm obronny przed pasożytami i chorobami u Apis mellifera. Jednak podstawa genomowa i ścieżki funkcjonalne zaangażowane w inicjację tego zachowania są nadal niejasne. Odszyfrowanie genomicznych podstaw zachowań higienicznych jest warunkiem wstępnym opracowania szerokiego repertuaru markerów genetycznych powiązanych z poziomem wydajności tej cechy.
Aby wypełnić tę lukę w wiedzy, przeprowadziliśmy sekwencję RNA na próbkach z mózgów 25 pszczół miodnych z pięciu uli higienicznych i trzech uli niehigienicznych. Analiza wykazała, że za zachowanie higieniczne pszczół miodnych wpływa konkretna liczba genów funkcjonalnych. Zidentyfikowane geny, a zwłaszcza ich lokalizacja w genomie pszczoły miodnej, są zgodne z wcześniejszymi ustaleniami.
Odkrycia te pogłębiają naszą wiedzę na temat uwarunkowań genetycznych zachowań higienicznych pszczół. Nasze wyniki pokazują, że zachowania higieniczne opierają się na konkretnym zestawie genów powiązanych z różnymi wzorcami regulacji, które z kolei są związane z nadekspresją genów kodujących cytochrom P450.
Zachowanie higieniczne u pszczół zostało po raz pierwszy opisane w 1937 roku przez Parka, ale jego podłoże genetyczne (że jest to instynkt kodowany wielogenowo) po raz pierwszy zasugerowali Rothenbuhler i in. (1964), który zaproponował model dwóch loci w celu wyjaśnienia dziedziczenia zachowań higienicznych. Od tego czasu zachowanie to uznawano za przykład wpływu genów odziedziczonych po mendlowsku. Uważano, że:
- jedno miejsce (u) bierze udział w odsklepianiu,
- a drugie (r) w usuwaniu zainfekowanej larwy.
- Homozygota jednego z loci powinna albo otworzyć czapeczkę (uu),
- albo usunąć larwę (rr). Później opracowano model trzech loci, aby lepiej dopasować oryginalne dane. Niedawno w czterech badaniach opartych na technikach molekularnych:
- RAPD - jedna z metod technimi PCR i,
https://experiments.springernature.com/articles/10.1385/1-59259-384-4:117
- SNP - Szukaniu i porównywaniu polimorfizmów pojedynczego nukleotydu – jest to zjawisko zmienności sekwencji DNA, która polega na zmianie pojedynczego nukleotydu pomiędzy osobnikami danego gatunku lub pomiędzy różnymi gatunkami w sekwencjach analogicznych. Pomiedz chromosomami homologicznymi w sekwencjach homogicznych.
- Stwierdzono odpowiednio siedem, sześć, dziewięć i dwa QTL - Quantitative Trait Loci (Locus cechy ilościowej). Metoda badań, stosowana między innymi w genetyce zachowania pozwala na poszukiwanie genów odpowiedzialnych za cechy warunkowane poligenowo. W tym przypadku powiązane z zachowaniami higienicznymi pszczół. Wyniki te sugerują, że podłoże genetyczne zachowań higienicznych jest bardziej złożone, niż wcześniej sądzono!
Próg wykrywania chorego czerwia zależy od tego, jak szybko robotnica-pielęgniarka może wykryć i zainicjować proces usuwania chorego czerwia. Wydaje się, że na tę detekcję wpływają zdolności węchowe pszczół karmiących. Ponadto wydaje się, że wszystkie pszczoły robotnice wykazują różny poziom zachowań higienicznych, a ich skuteczność jest powiązana z szybkością wykonania pełnego zadania (odsklepienie wieczka komory i usunięcie poczwarki)
Ekspresja genów w mózgu jest ściśle powiązana ze stanem zachowania pszczół miodnych. Dlatego, aby zapewnić identyfikację wiarygodnego sygnału korelującego zarówno ekspresję genów, jak i zachowania higieniczne zbadaliśmy tkankę mózgową pszczół karmiących z rodzin, które były najbardziej kontrastowe pod względem interesującej nas cechy fenotypowej.
W naszym badaniu przeanalizowaliśmy profile transkryptomiczne 13 hodowanych rodzin pszczół miodnych. Celem było zbadanie i porównanie zróżnicowanej ekspresji genów pomiędzy liniami higienicznymi i niehigienicznymi, aby zidentyfikować geny zaangażowane w zachowania higieniczne. Ostatecznie celem było dostarczenie funkcjonalnych markerów genetycznych do analizy SNP w celu opracowania narzędzi genomicznych przydatnych w programach selekcji pszczół miodnych pod kątem wykrywania szczepów higienicznych.
Spośród 11 168 genów występujących w genomie A. mellifera stwierdzono ekspresję 10 519 genów w pulach higienicznych i 10 374 genów w pulach niehigienicznych. 10 genów o największej ekspresji było takich samych dla dwóch zjawisk behawioralnych, ale ich kolejność była inna -geny zaangażowane w produkcję mleczka pszczelego.”
https://youtube.com/shorts/W_gHUyaFZOE?si=SawHi9kOPm0CaJIl
https://en.wikipedia.org/wiki/Epistasis
„Epistaza w genomach organizmów występuje w wyniku interakcji między genami. Ta interakcja może być bezpośrednia, jeśli geny kodują białka, które na przykład są oddzielnymi składnikami wieloskładnikowego kompleksu (takie jak rybosom wić bakteryjna, syntaza ATP..), wzajemnie hamują własną aktywność [sprzężenia dodatnie i ujemne], lub jeśli białko kodowane przez jeden gen modyfikuje drugi gen (np. przez fosforylację, albo niezbędną obróbkę potranskrypcyjną, potranslacyjną, modyfikowanie białek za pomocą proteaz - odcinanie sekwencji adresowych, żeby odsłonić sortujące, czy chaperonów ).
Alternatywnie taka interakcja może być pośrednia, w których geny kodują komponenty w szlaku metabolicznym, lub sieci szlaku rozwojowego, szlaku przekazywania sygnałów (transdukcji sygnału), lub kodujące czynniki transkrypcyjne, regulujące ekspresje danego genu w sieci (kolejność, czas wyrażania, co by się stało, gdyby geny kodujące białka rybosomowe nie ulegały precyzyjnej regulacji?). Na przykład gen kodujący enzym, który syntetyzuje penicylinę jest bezużyteczny dla grzyba bez enzymów, które syntetyzują niezbędne prekursory w szlaku metabolicznym.
[…..]
Wysoki poziom epistazy jest zwykle uważany za czynnik ograniczający ewolucję, a ulepszenia wysoce epistatycznej cechy są uważane za mające mniejszą zdolność ewolucji . Dzieje się tak, ponieważ na jakimkolwiek podłożu wielogenowym bardzo niewiele mutacji będzie korzystnych, (!) chociaż może zajść potrzeba wystąpienia wielu mutacji, aby ostatecznie poprawić cechę.”
https://youtube.com/shorts/ABtMIkWEgp4?si=DaIFPpmVD3kzn7pv
(!)
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2286568/

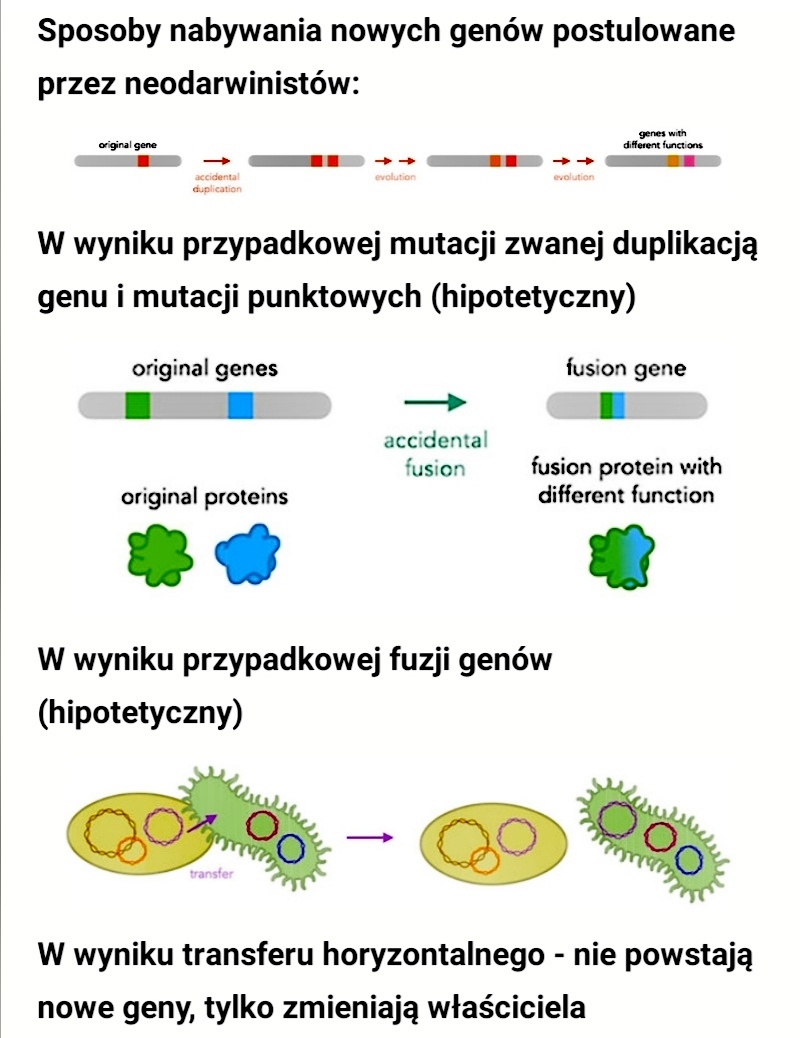
„Uważa się, że duplikacja genów jest głównym źródłem innowacji ewolucyjnych, ponieważ umożliwia jednej kopii genu mutację i eksplorację przestrzeni genetycznej, podczas gdy druga kopia nadal spełnia pierwotną funkcję.
Modele tego procesu często domyślnie zakładają, że pojedyncza mutacja w zduplikowanym genie może nadać nową wybieralną właściwość. Jednak niektóre cechy białek, takie jak wiązania dwusiarczkowe lub miejsca wiązania ligandów, wymagają udziału dwóch lub więcej reszt aminokwasowych, co może wymagać kilku mutacji. Tutaj modelujemy ewolucję takich cech białek za pomocą tego, co uważamy za najprostszą koncepcyjnie drogę - mutację punktową w zduplikowanych genach.
Pokazujemy, że dla bardzo dużych populacji N, gdzie w stanie ustalonym przy braku selekcji można oczekiwać, że populacja będzie zawierać jeden lub więcej zduplikowanych alleli kodujących użyteczną cechę, czas do utrwalenia się jej w populacji oscyluje w pobliżu odwrotności punktu współczynnika mutacji i zmienia się powoli wraz z λ^th z 1/N, gdzie λ to liczba pozycji nukleotydów, które muszą zostać zmutowane, aby wytworzyć daną cechę.
Przy mniejszych populacjach czas do utrwalenia zmienia się liniowo z 1/N i przekracza odwrotność częstości mutacji punktowych. Dochodzimy do wniosku, że ogólnie rzecz biorąc, aby określić w 10^8 pokoleniach, wytwarzanie nowych cech białek, które wymagają udziału dwóch lub więcej reszt aminokwasowych po prostu w wyniku wielokrotnych (mnogich, powstałych równocześnie) mutacji punktowych w zduplikowanych genach, wymagałoby populacji nie mniejszej niż 10^9.”
Wyobraź sobie proszę, że do powstania jakieś niezbędnej korzystnej funkcji potrzebne są aż 4 mutacje: A, B, C , D. Że dopiero zestaw A, B, C ,D może dać korzyść selekcyjną (dostosowanie). Jaka więc korzyść z pojedynczych mutacji: A, B , C czy D, skoro żadna z nich z osobna nie daje żadnej przewagi selekcyjnej? Każda z osobna jest neutralna, bezużyteczna? W tym przypadku, żeby cecha określana przez mutacje: A, B, C, D mogła dać przewagę selekcyjną, to te 4 [kompletny zestaw] mutacje musiałyby nastąpić za jednym zamachem.